Стебель хвойных растений
Строение молодого стебля сосны обыкновенной (Pinus sylvestris L.)
Для изучения анатомического строения стебля могут быть использованы 3—4-летние удлиненные побеги сосны, выросшей в благоприятных условиях освещения. Лучше всего собирать материал во второй половине лета, когда прирост побегов в длину закончен. Материал фиксируют в спирте.
Поперечные срезы последовательно обрабатывают флороглюцином, соляной кислотой и заключают в глицерин. До проведения реакции на одревеснение срезы можно обработать раствором иода в водном растворе йодистого калия. Для работы пригодны срезы небольших размеров; важно только, чтобы в них были и периферические, и внутренние участки стебля, включающие сердцевину. Анатомическую топографию стебля изучают при малом увеличении, некоторые детали строения— при большом увеличении микроскопа.
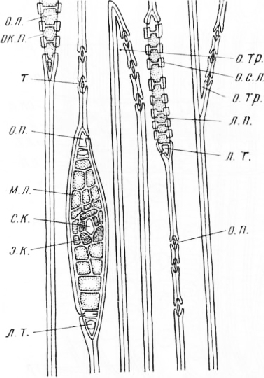
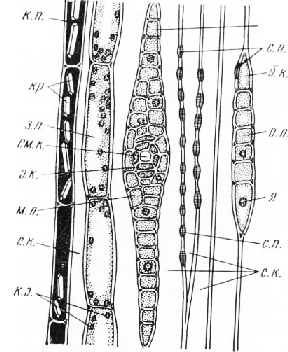
Стебель сосны в поперечном сечении более или менее округлый.
Вокруг нее расположена слабо развитая первичная древесина, элементы которой нередко оказываются более одревесневшими, чем примыкающие к ним снаружи клетки вторичной древесины. Во вторичной древесине можно различить кольца прироста. Древесина состоит из толстостенных трахеид, расположенных радиальными рядами. В каждом кольце прироста видны схизогенные смоляные каналы, окруженные обкладкой из неодревесневших тонкостенных паренхимных клеток. Радиальные ряды трахеид последнего, самого молодого кольца прироста постепенно переходят в камбиальную зону, состоящую из нескольких слоев тонкостенных узко-просветных клеток, вытянутых в тангентальном направлении.

Вокруг камбиальной зоны узким кольцом расположен вторичный луб, или вторичная флоэма. Первичная флоэма, находящаяся на границе с первичной корой, практически неразличима.
Вторичный луб состоит из очень мелких светлых таблитчатых в поперечном сечении ситовидных элементов и более или менее округлых паренхимных клеток, составляющих короткие тангентальные цепочки. На срезе обработанном раствором иода в водном растворе йодистого калия, в паренхимных клетках можно обнаружить запасной крахмал. Все элементы луба расположены радиальными рядами.
В радиальном направлении древесина и луб пересечены узкими, обычно однорядными лучами разной протяженности. Формирование лучей начинается в период первичной дифференцировки анатомической структуры стебля, когда центральный цилиндр имеет еще пучковое строение. При вторичном утолщении нарастание лучей в радиальном направлении осуществляется за счет камбия, производные которого раздвигают концевые участки лучей, расположенные в зонах хорошо сохраняющейся в стеблях первичной древесины и плохо заметной первичной флоэмы. Хотя в стеблях с вторичным утолщением почти весь луч составлен клетками вторичного (камбиального) происхождения, за этими лучами можно сохранить название первичных, как это было принято в старой учебной анатомической литературе. Первичные лучи располагаются между первичной корой и сердцевиной. Этим, вероятно, и объясняется также то, что в русской и зарубежной литературе лучи называют сердцевинными, применяя этот термин и к лучам вторичного происхождения, возникающим в любом возрасте побега вследствие деления коротких камбиальных клеток. Вторичные лучи по сравнению с первичными имеют меньшую протяженность в радиальном направлении, никогда не доходят до первичной коры и сердцевины, но по строению не отличаются от первичных лучей.
В лубе лучи строго радиальны лишь вблизи камбиальной зоны, по мере удаления от нее они слегка искривляются. Некоторые лучи на конце расширены и вдаются в первичную кору. Расширения представляют собой замкнутые смоловместилища шаровидной или мешковидной формы.
Первичная кора довольно узкая, наружная часть ее состоит из клеток, более или менее вытянутых в тангентальном направлении. В первичной коре хорошо заметны округлые или овальные в поперечном сечении вертикальные смоляные каналы, расположенные вокруг центрального цилиндра.
Первичная кора окружена перидермой, все элементы которой расположены радиальными рядами. Наружная часть перидермы состоит из нескольких слоев клеток пробки, внутренняя — из феллогена и трех-четы-рех слоев клеток феллодермы. Снаружи от перидермы видны сухие, легко отрывающиеся от стебля чешуйки, представляющие собой остатки отмершего эпидермиса и нескольких наружных слоев клеток первичной коры.
Задание. При малом увеличении микроскопа зарисовать схему строения многолетней ветки, отметив сердцевину, первичную древесину, вторичную древесину с кольцами прироста, камбиальную зону, луб, лубо-древесин-ные (сердцевинные) лучи, первичную кору со смоляными каналами и перидерму.
Строение проводящих тканей
Для детального изучения строения проводящих тканей—древесины и луба (флоэмы)—лучше использовать древесину и кору стволов взрослых (не моложе 10—15 лет) деревьев, так как в стволе структурные особенности элементов, слагающих проводящую систему, выражены лучше, чем в ветвях.
Древесина и луб представляют собой сложные ткани, состоящие из элементов, различных по строению, функции и расположению. Такие ткани изучают на срезах, проведенных в трех взаимно перпендикулярных направлениях: поперечном, перпендикулярном продольной оси органа, продольном радиальном, плоскость которого проходит по радиусу перпендикулярно кольцам прироста древесины, и продольном, тангентальном, проведенном по касательной к ним, перпендикулярно сердцевинным лучам (рис. 1).

Небольшие кусочки стволовой коры снимают вместе с камбиальной зоной и периферическим кольцом древесины Место, с которого снята кора, следует замазать садовым варом. Материал собирают летом до окончания вегетационного периода и фиксируют в спирте.
Кусочки древесины хранят в спирте с глицерином. Перед изготовлением срезов древесину для размягчения кипятят в воде в течение 1—2 ч и режут в горячем состоянии, смазывая глицерином поверхность среза, которую предварительно выравнивают скальпелем, ориентируясь по кольцам прироста. При изготовлении поперечных срезов бритву следует вести не поперек, а вдоль этих колец. В таком направлении древесину резать легче. Срез должен захватить два-три кольца прироста. Для получения продольных срезов необходимо сделать небольшие поперечные зарубки на расстоянии 5—7 мм от торца образца. Это предохранит от поранения, если бритва сорвется.
Строение древесины сосны обыкновенной (Pinus sylvestris L.)
Древесина состоит из продольных трахеид, обеспечивающих восходящий ток воды и растворенных в ней минеральных веществ, а также выполняющих механическую функцию; лучевых трахеид, обусловливающих перемещение водных растворов в горизонтальном (радиальном) направлении; паренхимных клеток, входящих в состав лучей и обкладки смоляных каналов; эпителия, выстилающего полость канала и участвующего в выделении смол, эфирных масел и бальзамов.
Срезы древесины рассматривают в глицерине.
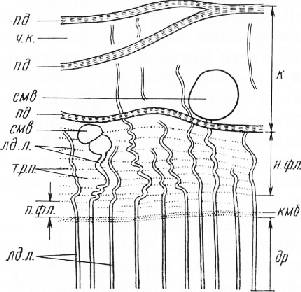
На поперечном срезе хорошо выражены кольца прироста (рис. 48). Внутренняя часть каждого кольца состоит из довольно широкопросветных, в очертании многоугольных трахеид, слагающих раннюю, или весеннюю, древесину. Периферическая часть кольца прироста, образующаяся во второй половине вегетационного периода, представлена более толстостенными уз-копросветными трахеидами, составляющими позднюю, или летнюю, древесину. Обычно она темнее ранней.
В пределах каждого кольца переход от ранней древесины к поздней постепенный. Трахеиды расположены радиальными рядами. Весенние (ранние) трахеиды выполняют главным образом функцию проведения водных растворов. На их радиальных стенках видны крупные окаймленные поры. Они отличаются от простых пор тем, что вторичные оболочки трахеид, образующие стенки камеры поры, сводообразно приподняты над замыкающей пленкой, или мембраной поры. В середине свода находится отверстие, соединяющее камеру поры с полостью трахеиды.

Если разрез прошел через боковые части свода, то пора выглядит как линзовидное расширение клеточной стенки. Стенки трахеид летней (поздней) древесины имеют небольшое число пор. Основная функция этих трахеид —механическая. Между радиальными рядами трахеид расположены древесинные лучи, состоящие обычно из одного ряда клеток, структурные особенности которых лучше рассматривать на радиальном срезе.
Смолов ы делительная система древесины представлена вертикальными и горизонтальными смоляными канала-м и. Вертикальные смоляные каналы обычно расположены в поздней древесине кольца прироста. На срезе они видны в поперечном сечении. Канал окружен обкладкой из паренхимных клеток. Полость канала выстлана тонкостенными эпителиальными клетками. При изготовлении среза нежные эпителиальные Радиальный
клетки нередко разрушаются, срез древесины дает представление о трахеидах как очень длинных, тых в продольном направлении прозенхимных клетках со слегка закругленными окончаниями. Между концами трахеид, расположенными на одном уровне, ‘вклиниваются трахеиды, принадлежащие другому ярусу.
Окаймленные поры, находящиеся на радиальных тенках трахеид, видны в плане. Они представляют со-ои совокупность трех концентрических окружностей, самая внутренняя из которых соответствует отверстию камеры поры, средняя — очертаниям торуса, внешняя — контурам окаймления.

Древесинные лучи на радиальном срезе имеют вид широких горизонтальных полос, состоящих из нескольких рядов клеток. Внутренняя часть луча сложена тонкостенными клетками лучевой паренхимы, вытянутыми в радиальном направлении. Клетки содержат крахмал и капли масла. В старой древесине протопласты клеток лучевой паренхимы отмирают. В месте пересечения с каждой продольной трахеи-дой в стенке паренхимной клетки формируется по одной крупной округло-четырехугольной простой поре. Эти поры называют оконцевыми или ок-новидными. Общее число пор в клетке зависит от числа трахеид, с которыми данная клетка соприкасается. Со стороны трахеиды поры имеют небольшое окаймление. Такие полуокаймленные поры можно было видеть на поперечном срезе древесины, если его плоскость прошла через середину луча. По краям луча располагаются лучевые трахеиды, или трахеидальные клетки. Это мертвые клетки, обеспечивающие перемещение воды в радиальном направлении. Обычно они короче клеток лучевой паренхимы и часто имеют неправильные очертания. Внутренняя поверхность их клеточных стенок зубчатая. Многочисленные мелкие окаймленные поры расположены на всех стенках. Лучи, состоящие из клеток функционально и морфологически неоднородных, называют гетерогенными.
На тангентальном срезе вертикальные трахеиды имеют заостренные окончания. Поры на их радиальных стенках видны в разрезе. Древесинные лучи вклинивающиеся между трахеидами, перерезаны поперек. Они представляют собой цепочки коротких округлых или квадратно-прямоугольных клеток. Средняя часть луча занята клетками запасающей паренхимы, краевые клетки луча представлены лучевыми трахеидами. Наряду с однорядными лучами высотой 1 —12 клеток встречаются сложные многорядные веретеновидные лучи, в расширенной средней части которых находится по одному смоляному каналу, принадлежащему горизонтальной смоловыделительной системе. На продольных срезах могут быть и вертикальные смоляные каналы в виде широких полос, состоящих либо из эпителиальных клеток, либо из клеток паренхимной обкладки. Если срез пройдет через середину полости канала, то срез распадется на две части.
Задание.
1. При большом увеличении микроскопа зарисовать участок поперечного среза на границе двух колец прироста, обратив внимание на характер поперечного сечения трахеид ранней и поздней древесины; отметить поры на радиальных стенках, изобразить древесинный луч и смоляной канал в поздней древесине.
2. Зарисовать участок радиального среза, отметив особенности строения продольных трахеид с окаймленными порами, показать луч, состоящий из лучевых трахеид с мелкими окаймленными порами и клеток паренхимы с порами в стенках, смежных со стенками продольных трахеид.
3. Зарисовать строение древесины на тангентальном срезе, отметив трахеиды с цепочками окаймленных пор на перерезанных радиальных стенках; изобразить простои, однорядный, и сложный, многорядный, лучи.
Строение вторичного луба сосны обыкновенной (Pinus sylvestris L.)
Вторичный луб (флоэму), так же как и древесину, изучают на поперечных, радиальных и тангентальных срезах.
В проведении продуктов ассимиляции участвует лишь самая молодая часть луба, прилегающая к камбию. Ширина ее обычно не превышает 0,3 мм. Снаружи от нее расположена более старая и более широкая зона, в которой проводящие элементы уже не функционируют. Эту зону называют непроводящей. У сосны непроводящий луб выполняет функции запаса питательных веществ и выделения. Луб окружен чешуйчатой коркой (см. с. 143).
Вторичный луб, или флоэма, состоит из ситовидных элементов, проводящих растворы органических соединений, продольных тяжей паренхимных клеток, лучевой паренхимы и смоловыделитель-ных эпителиальных клеток.
Функциональные различия между проводящей и непроводящей зонами луба коррелируют с их структурными особенностями.
На поперечном срезе элементы проводящей зоны расположены радиальными рядами. Ситовидные клетки тонкостенные, широкопросвет-ные, квадратных или прямоугольных очертаний.
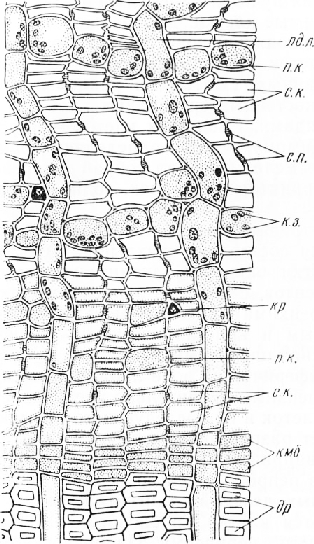
Лучи преимущественно однорядные, реже состоят из нескольких рядов вытянутых в радиальном направлении паренхимных клеток с цитоплазмой, ядром, иногда с зернами крахмала. В проводящей зоне лучи строго радиальны, в непроводящей они извилисты, некоторые лучи местами расширены. Мешковидные или головчатые расширения лучей представляют собой замкнутые смо-ловместилища. Они формируются в результате преобразования лучевой паренхимы в эпителиальные смоло-выделительные клетки.
В непроводящей зоне слои широкопросветных ситовидных клеток обычно чередуются со слоями сильно деформированных, сжатых в радиальном направлении очень тонкостенных ситовидных клеток, расположенных косыми рядами. Слои этих клеток, дифференцирующихся, по-видимому, весной, определяют границы годичных приростов. Если материал собран с деревьев, выросших в неблагоприятных условиях, годичная слоистость луба не видна.
Во внутренних частях непроводящей зоны паренхим-ные клетки составляют рыхлые тангентальные цепочки. Одни клетки округлые, значительно крупнее проводящих элементов, с зернами запасного крахмала (запасающая паренхима), другие — мелкие, угловатые, с бурым содержимым и кристаллическими включениями (кри-сталлоносная паренхима).
В периферической зоне луба, примыкающей к корке, объем паренхимы значительно увеличивается вследствие деления клеток и их последующего растяжения. Это обусловливает разрастание (дилатацию) луба в танген-тальном направлении. Из паренхимных клеток наружных слоев непроводящей зоны в результате двух последовательных делений тангентальными перегородками вычленяются клетки пробкового камбия (феллогена), образующего перидерму. На поперечном срезе внутренние перидермы располагаются отдельными дугами.
На продольном радиальном срезе видно, что ситовидные элементы по строению сходны с трахеидами. Это очень длинные клетки с притуплёнными концами. На их радиальных стенках расположены в один ряд округлые ситовидные поля с очень мелкими прободениями. На срезе, включающем проводящий луб, камбиальную зону и древесину, можно проследить постепенное формирование ситовидных полей, которые обычно находятся почти на одном уровне с окаймленными порами в трахеидах.
Паренхимные клетки, находящиеся между ситовидными элементами, вытянуты в продольном направлении. Они составляют тяжи, конечные клетки которых имеют клиновидные очертания. Такую паренхиму называют тяжевой. В проводящей зоне паренхимные клетки по ширине почти не отличаются от ситовидных элементов, в непроводящей — они более широкие. На периферии этой зоны правильность их расположения продольными тяжами постепенно исчезает, клетки округляются, между ними возникают межклетники. Кроме клеток запасающей паренхимы в тяже могут быть и кристаллоносные клетки с одиночными призматическими кристаллами оксалата кальция, окруженными матриксом из дубильных веществ, смол и других соединений, обусловливающих бурую окраску содержимого этих клеток. Кристаллоносные клетки обычно мертвые.
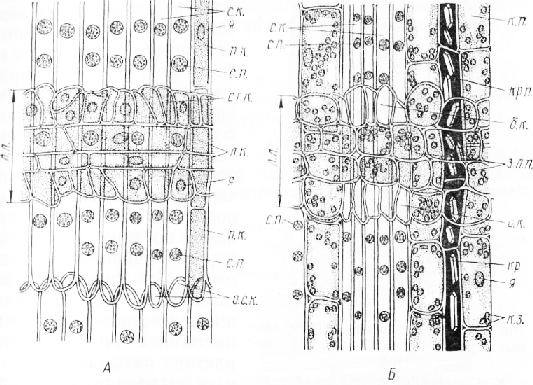
Лубяные лучи представляют собой горизонтальные полосы, состоящие из нескольких рядов клеток.
На срезах непроводящего луба лучи часто прерывисты, так как в этой зоне они проходят не строго ради-ально. Лучи в лубе сосны гетерогенные. Внутренние их клетки, вытянутые в радиальном направлении, называют лежачими. В них можно видеть цитоплазму и ядро,
в непроводящем лубе эти -ил клетки содержат запасной крахмал. Краевые клетки, вытянутые в вертикальном направлении, называют стоячими. Те из них, которые соприкасаются с тяжевой паренхимой, выполняют функцию запаса, а клетки, контактирующие с ситовидными элементами через мелкие ситовидные поля, имеют цитоплазму, ядро, но никогда не содержат запасного крахмала. По функции они сходны с сопровождающими клетками флоэмы цветковых растений. Эти клетки называют «белковыми» или клетками Страсбургера. «Белковые» клетки после прекращения деятельности ситовидных элементов отмирают, поэтому в непроводящем лубе они очень мелкие, деформированные и выглядят пустыми в отличие от крупных округлых запасающих клеток лучевой паренхимы. Если срез прошел через смоловместилище, то внутри луча можно видеть полость, окруженную многочисленными тонкостенными эпителиальными клетками.
Тангентальный срез проводящего луба расположением элементов напоминает тангентальный срез древесины. На радиальных стенках ситовидных элементов ситовидные поля видны в разрезе. Хорошо выражена тяжевая паренхима, состоящая из крахмалоносных и кристаллоносных клеток. Лубяные лучи двух типов: однорядные, сложенные округлыми или округло-четырехугольными клетками, и более высокие много-рядные с перерезанным поперек смоляным каналом, который представляет собой продолжение горизонтального смоляного канала древесины. В непроводящей зоне луба смоляные каналы расширяются, увеличивается число выстилающих их эпителиальных клеток. Остальные клетки луча неравномерно разрастаются, и некоторые из них по внешнему виду почти не отличаются от клеток тяжевой паренхимы.
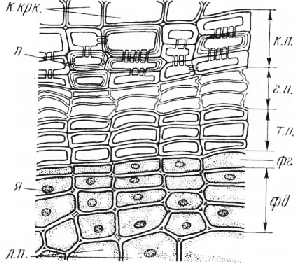
Строение перидермы и корки сосны обыкновенной (Pinus sylvestris L.)
Стволы 10—15-летних и более старых деревьев имеют корку — комплекс мертвых наружных участков луба, разделенных перидермами, которые на поперечных срезах имеют вид дуг. Такую корку называют чешуйчатой.
Перидермы, разделяющие чешуи корки, состоят из нескольких слоев клеток, расположенных радиальными рядами. Три-четыре самых внутренних слоя представляют собой фел-лодерму. В самой внутренней перидерме, расположенной на границе с жизнедеятельным лубом, клетки феллодермы живые, со слегка утолщенными
стенками. Кнаружи от нее располагается феллоген, или пробковый камбий, в виде однорядного слоя таблитчатых клеток. В наружных перидермах, в которых деятельность феллогена прекращена, его клетки неотличимы от клеток феллодермы. Периферические клетки перидермы составляют пробку, или феллему. У сосны она состоит из трех типов клеток. Внутренние клетки имеют слегка утолщенные стенки, содержащие суберин. Это типичные клетки пробки. К ним примыкают прозрачные клетки с очень тонкими стенками. Радиальные стенки их обычно извилисты. Это так называемая губчатая пробка. Наружные слои перидермы представлены клетками с очень толстыми одревесневшими слоистыми стенками, в которых обычно хорошо видны многочисленные узкие норовые каналы. Эти клетки по строению сходны с каменистыми клетками. Их называют феллоидами. Так как связь между клетками-феллоидами и губчатой пробкой очень непрочна, между ними легко возникают разрывы, способствующие опадению чешуй корки.
Задание.
1. При малом увеличении микроскопа зарисовать схему строения вторичного луба и корки, отметив проводящую и непроводящую зоны, однорядные лучи и лучи со смоловместилищами.
2. При большом увеличении микроскопа зарисовать участки поперечного, радиального и тангентального срезов, показав на рисунках расположение и общий вид ситовидных клеток с ситовидными полями на радиальных стенках, клетки крахмалоносной и кристаллоносной тяжевой паренхимы; на радиальном срезе непроводящей зоны луба обратить внимание на строение луча, отметить в нем отмершие «белковые» клетки и клетки запасающей паренхимы.
3. При большом увеличении микроскопа зарисовать три типа клеток пробки.
- * *
Вместо сосны со строением стебля хвойных растений можно ознакомиться на примере лиственницы (Larix) или ели (Picea).
В молодых 3—4-летних стеблях лиственницы вертикальных смоляных каналов в первичной коре может не быть, так как содержащая их наружная часть первичной коры рано сбрасывается вследствие более глубокого, чем у сосны, заложения феллогена и образования перидермы. Древесина лиственницы и ели отличается от древесины сосны толстостенными эпителиальными клетками, выстилающими полости смоляных каналов. Поры на радиальных стенках трахеид лиственницы иногда располагаются в два ряда. На смежных стенках между вертикальными трахеидами и паренхимными клетками образуются не оконцевые, а мелкие окаймленные поры, у лиственницы иногда встречается тяжевая древесинная паренхима.
В непроводящей зоне вторичной флоэмы этих деревьев наряду с ситовидными клетками, запасающей и кри-гтяллоносной папенхимой имеются механические элементы. У ели это группы каменистых клеток, у лиственницы — волокнистые склереиды — длинные прозенхимные толстостенные клетки, которые на поперечном срезе имеют округлые очертания.
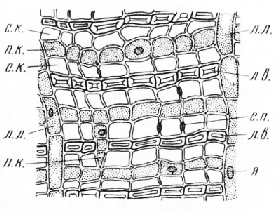
Интересно строение древесины и луба тиса (Taxus), кипариса (Сир-ressus) и можжевельника (Juniperus). В их древесине нет смоляных каналов, но хорошо развита тяжевая паренхима. Тра-хеиды тиса кроме пор имеют спиральные утолщения, образующиеся, видимо, за счет третичной оболочки. Древесинные лучи у этих пород сложены однородными клетками. Такие лучи называют гомогенными.
У кипариса и можжевельника луб состоит из чередующихся тангентальных однорядных слоев проводящих, паренхимных и механических элементов, которые в свою очередь располагаются радиальными рядами. Больших различий в строении проводящей и непроводящей зон луба нет. На поперечном срезе в радиальном ряду между двумя механическими элементами, представляющими собой очень длинные волокна с одревесневшими стенками, расположены примыкающие к ним ситовидные клетки, разделенные одной, очень редко двумя паренхимными клетками. Кристаллоносной паренхимы в лубе этих древесных пород нет. В ситовидных клетках после прекращения их деятельности откладывается мелкий кристаллический песок.
Анатомия растений - Стебель хвойных растений