Стебель травянистых двудольных растений
Стебель кирказона крупнолистного (Aristolochia macrophylla Lam.)
Кирказон (лиана, которую часто разводят в садах) — один из лучших объектов для изучения строения стебля так называемого пучкового типа.
Для этой цели используют стебли однолетнего вегетативного побега, образовавшегося в текущем году. Материал фиксируют спиртом в конце вегетационного периода. Лучше всего вырезать кусочки стеблей из междоузлий, расположенных в средней части однолетнего побега, у которых закончилась первичная дифференцировка анатомической структуры, а вторичных тканей еще не очень много.
Строение стебля изучают на тонких поперечных срезах, которые могут быть неполными, однако необходимо, чтобы они захватили все ткани от сердцевины до эпидермиса.
После предварительного рассмотрения в капле воды с целью установления пригодности для изучения срезы обрабатывают спиртовым раствором флороглюцина, затем соляной кислотой и заключают в глицерин. Можно сначала поместить срез в раствор иода в водном растворе иодистого калия, а потом провести реакцию на одревеснение.
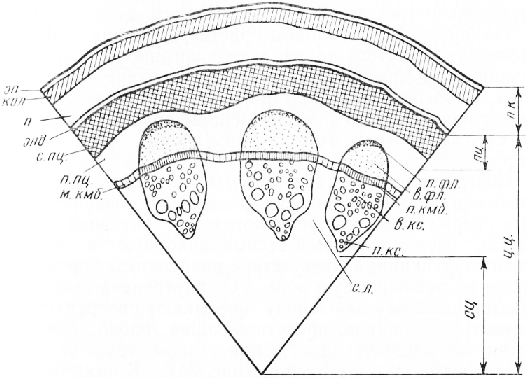
На поперечном срезе (рис. 33) при малом увеличении (56—80 раз) можно различить: 1) эпидермис; 2) первичную кору, состоящую из нескольких слоев клеток, и 3) центральный цилиндр, или стель. Центральный цилиндр начинается широким кольцом склеренхимы , состоящей из толстостенных клеток с одревесневшими оболочками, которые после проведения реакции на одревеснение приобрели вишнево-красный цвет. Наружная граница склеренхимного кольца ровная, внутренняя— волнистая, с небольшими углублениями против проводящих пучков. Кольцо склеренхимы вместе с подстилающим его довольно широким слоем паренхимы составляет перицикл. Ковнутри от него расположен один круг проводящих пучков, ксилемная часть которых тоже приобрела красный цвет.
Флоэма отличается от паренхимы перицикла более желтой окраской и мелкими размерами клеток. Пучки разделены сердцевинными лучами, состоящими из паренхимных клеток. Центральную часть стебля занимает широкая сердцевина.
Ознакомившись с общей топографией среза, следует перейти к изучению слагающих его тканей при большом (400—600 раз) увеличении микроскопа.
Эпидермис состоит из почти прямоугольных плотно сомкнутых клеток. Их наружные стенки значительно толще боковых и внутренних. С поверхности эпидермис покрыт сплошным тонким слоем кутикулы, которая усиливает его защитную роль.
Под эпидермисом в наружной части первичной коры расположены несколько слоев клеток пластинчатой или, реже, уголков ой колленхимы. Клетки ее слегка вытянуты в тангентальном направлении.
Глубже колленхимы лежат крупные тонкостенные па-ренхимные клетки. Присутствие хлоропластов в парен-химных клетках и элементах колленхимы обусловливает зеленую окраску молодых участков стебля. До проведения реакции па одревеснение в некоторых паренхимных клетках можно видеть друзы щавелевокислого кальция, растворяющегося потом от действия соляной кислоты.
На срезе, обработанном раствором иода в водном растворе йодистого калия, в паренхимных клетках первичной коры можно обнаружить зерна запасного крахмала. Особенно много крахмала в клетках самого внутреннего однорядного слоя первичной коры, составляющего крахмалоносное влагалище, или эндодерм у.
Содержание в клетках зеленых пластид, запасного крахмала и кристаллических включений позволяет считать первичную кору физиологически активной зоной стебля, осуществляющей ассимиляционную и выделительную функции, а присутствие колленхимы определяет также ее механическую роль. С внутренней стороны к крахмалоносному влагалищу примыкают склеренхимные элементы перицикла, представляющие собой волокна, вытянутые вдоль стебля. На поперечном срезе они имеют многоугольные очертания. Клетки плотно соединены между собой, оболочки их толстые, одревесневшие. Наиболее сильно одревесневают оболочки волокон, составляющих периферическую часть перицикла. Богатые лигнином первичные оболочки и межклеточные пластинки на срезах после проведения реакции на одревеснение окрашены темнее, чем вторичные оболочки, имеющие меньше лигнина.
Внутренняя зона перицикла состоит из крупных живых тонкостенных паренхимных клеток.
Проводящие пучки, расположенные кольцом внутрь от перицикла, в очертании округлые или овальные, слегка вытянутые в радиальном направлении. Пучки коллатеральные, открытые. Флоэма в пучке занимает наружное, ксилема — внутреннее положение. Между ними находится камбиальная зона из тонкостенных таблитчатых клеток, составляющих радиальные ряды.
Большая часть флоэмы образована камбием. Это вторичная флоэма. Ситовидные трубки флоэмы на срезе наиболее широкопросветны, вплотную к ним примыкают сопровождающие клетки, отличающиеся от паренхимных элементов флоэмы мелкими размерами и более темным содержимым. Первичная флоэма занимает самую периферическую часть пучка и состоит из тонкостенных, сильно деформированных клеток.
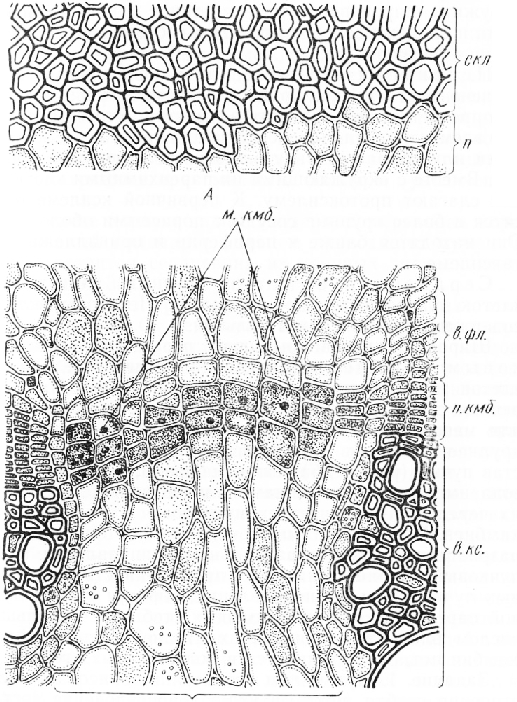
Вторичная ксилема, образованная камбием и расположенная внутрь от камбиальной зоны, состоит из крупнопросветных сосудов, значительного числа волокон и паренхимных клеток. Оболочки всех элементов ксилемы одревесневают. Изредка вблизи камбия можно обнаружить широкопросветные клетки с тонкими оболочками и следами содержимого. Это сосуды, еще не закончившие развитие.
Первичная ксилема, занимающая самую внутреннюю часть пучка, представлена небольшим числом узкопросветных проводящих элементов. Это самые ранние элементы ксилемы — сосуды или трахеиды с кольчатыми или спиральными утолщениями вторичных оболочек. Вместе с окружающими их паренхимными клетками они слагают протоксилему. К первичной ксилеме относятся и более крупные сосуды с пористыми оболочками. Они находятся ближе к периферии и принадлежат ме-таксилеме.
Сердцевина состоит из крупных паренхимных клеток, сложенных довольно рыхло. Некоторые клетки содержат друзы оксалата кальция.
Широкие сердцевинные лучи пересечены меж пучковым камбием, возникающим вследствие делений клеток луча тангентальными перегородками. На поперечном срезе клетки межпучкового камбия имеют более или менее прямоугольные очертания. Они значительно крупнее элементов камбиальной зоны, входящей в состав пучка. Таким образом, в стебле кирка-зсна имеется сплошное камбиальное кольцо, состоящее из чередующихся участков пучкового и межпучкового камбиев. Деятельность пучкового камбия обусловливает разрастание пучков в радиальном направлении. Межпучковый камбий увеличивает протяженность сердцевинных лучей в том же направлении, образуя клетки лучевой паренхимы. В очень молодых стеблях с небольшим числом элементов проводящих тканей межпучкового камбия нет, он формируется в более старых стеблях.
Задание.
1. При малом увеличении зарисовать схему строения стебля, показывающую распределение составляющих его тканей. На схеме отметить зону первичной коры и центрального цилиндра.
2. При большом увеличении микроскопа зарисовать клетки склеренхимы перицикла, пучкового и межпучкового камбиев.
Стебель подсолнечника однолетнего (Helianthus annuus L.)
Примером строения стебля пучкового типа с межпучковым камбием может служить также стебель подсолнечника.
Фиксированные в спирте кусочки стебля толщиной 0,5—0,7 см, вырезанные из междоузлий, разрезают вдоль по диаметру и делают поперечный срез, который должен захватить периферическую, плотную, и внутреннюю, более рыхлую, зоны стебля. После обработки срезов раствором иода в водном растворе йодистого калия проводят реакцию на одревеснение. Срезы заключают в глицерин и рассматривают при малом и большом увеличениях микроскопа.
На срезе хорошо видны эпидермис, первичная кора, центральный цилиндр с кольцом проводящих пучков и мощно развитая сердцевина из крупных паренхимных клеток.
Клетки эпидермиса имеют типичное для этой ткани строение. От некоторых клеток отходят многоклеточные волоски, состоящие из одного ряда клеток. Клетки более крупных прямых волосков имеют утолщенные оболочки и заканчиваются острием. В основании такого волоска лежит небольшая группа клеток, приподнимающая волосок над эпидермисом. Эти волоски придают стеблю шероховатость. Более мелкие, изогнутые волоски состоят из клеток с тонкими оболочками.
Под эпидермисом расположены несколько слоев клеток колленхимы. В молодых стеблях она уголковая, в более старых переходит в пластинчатую. На срезах, обработанных раствором иода в водном растворе йодистого калия, в клетках колленхимы заметны посиневшие или почти черные зерна крахмала, которые видны и в глубже лежащих тонкостенных клетках паренхимы. Особенно много крахмала в эндодерме, граничащей с наружной частью центрального цилиндра и выполняющей в стебле функцию крахмалоносного влагалища. В старых стеблях на радиальных стенках клеток эндодермы иногда видны пятна Каспари, которые дают положительную реакцию на одревеснение. Колленхима и паренхима, заканчивающиеся эндодермой, составляют первичную кору. В ней довольно много вместилищ выделений. Вместилище представляет собой длинный, в виде канала, межклетник, возникающий вследствие расхождения первоначально плотно сомкнутых клеток. Такие межклетники называют схизоген-н ы м и (от греч. «схизео» —- раздвигать) в отличие от лизигенных, формирование которых обусловлено растворением клеток (от греч. «лизис»—растворение). Клетки, окружающие канал, обычно делятся перегородками, параллельными очертаниям его полости. Внут-пенние из образовавшихся клеток составляют эпителий, Оделяющий в полость канала смолистые вещества.
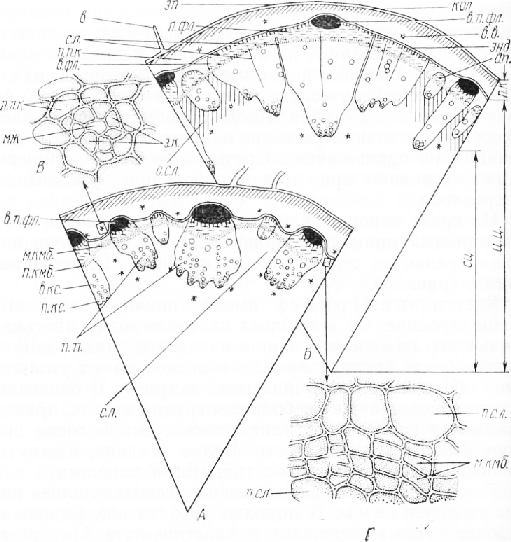
На рассматриваемом срезе вместилище видно в поперечном сечении. Вокруг межклетника расположен один ряд мелких эпителиальных клеток с густым содержимым. Полость межклетника обычно пустая, так как вещества, выделяемые эпителиальными клетками, растворяются спиртом при фиксации стебля. Схизогенные вместилища (рис. 3, В) встречаются в паренхиме центрального цилиндра, наружную часть которого составляет кольцо открытых коллатеральных проводящих пучков. Перицикла в стебле подсолнечника нет. В крупных пучках снаружи располагаются тяжи одревесневших толстостенных механических элементов, которые играют роль арматуры, предохраняющей глубже лежащие тонкостенные клетки флоэмы от деформации при изгибе стебля. Такие пучки называют сосудисто-волокнистыми или армированными. Механические элементы представляют собой волокна прокамбиального происхождения, относящиеся к протофлоэме. В мелких пучках арматуры нет.
Флоэма сложена многоугольными клетками разной величины. Более крупные из них — ситовидные трубки. Если срез захватил окончания члеников трубки, то можно увидеть пластинку с ситовидными прободениями. Если срез прошел через середину трубки, рядом с ней можно видеть мелкие сопровождающие клетки с густым зернистым содержимым. Сопровождающие клетки мельче паренхимных элементов флоэмы.
Расположенная внутрь от флоэмы камбиальная зона представляет собой широкую полосу мелких, расположенных радиальными рядами тонкостенных клеток. Все клетки каждого из этих рядов возникают в результате деления одной собственно камбиальной клетки и дифференцируются впоследствии в элементы ксилемы или флоэмы. Значительная ширина камбиальной зоны в молодом стебле свидетельствует об энергичном делении клеток камбия.
Ксилема, слагающая внутреннюю часть пучка, на срезах имеет разные очертания. Узкопросветные, округлые в поперечном сечении кольчатые и спиральные сосуды протоксилемы, расположенные близ сердцевины, а также наиболее ранние проводящие элементы метакси-лемы составляют радиальные цепочки, разделенные живыми тонкостенными паренхимными клетками.
В состав вторичной ксилемы, примыкающей к камбию, входят сосуды, древесинные волокна (либриформ) и паренхимные клетки. Наиболее широкопросветные элементы ксилемы — сосуды с многочисленными порами на боковых стенках. На поперечных срезах хорошо видны окаймления пор. Волокна либриформа в поперечном сечении имеют прямоугольные или многоугольные очертания. В клетках нет содержимого, оболочки их утолщенные, одревесневшие. Паренхимные клетки тонкостенные, нередко в них имеется содержимое.
Проводящие пучки разделены широкими сердцевинными лучами из паренхимных клеток.
Центральная часть стебля занята широкой сердцевиной. Клетки сердцевины паренхимные, тонкостенные, между ними обычно образуются межклетники. На ранних этапах развития стебля оболочки клеток неодревесневшие, прозрачные. В периферической части сердцевины клетки более мелкие, чем во внутренней.
Общий вид центрального цилиндра зависит от возраста стебля. Степень развития проводящих тканей в разных пучках различна, поэтому пучки. неодинаковы по размерам. Это объясняется тем, что конус нарастания побега подсолнечника долго сохраняет способность к формированию зачатков листьев, при этом из каждого зачатка в образовательное кольцо стебля входит по три прокамбиальных тяжа, дифференцирующихся неодновременно и различающихся по толщине. В разное время начинается в них развитие элементов первичных проводящих тканей и вторичное утолщение. В очень молодых стеблях между более или менее сформированными пучками могут быть и мелкие тяжи клеток прокамбия.
На более поздних этапах развития побега в связи с заложением новых листьев между пучками на уровне пучкового камбия из паренхимных клеток сердцевинных лучей вследствие их деления тангентальными перегородками вычленяются элементы межпучкового камбия. Таким образом в стебле развивается сплошное камбиальное кольцо.
Из межпучкового камбия возникает не только паренхима сердцевинных лучей, в нем дифференцируется также пучковый камбий, который образует элементы вторичных проводящих тканей, составляющих добавочные пучки. От ранее сформированных пучков они от-тичаются отсутствием радиальных цепочек сосудов про-токсилемы и арматуры над флоэмой. Участие межпучко-вого камбия в формировании добавочных пучков увеличивает мощность проводящей системы. Вследствие образования этих пучков число сердцевинных лучей увеличивается, но лучи становятся более узкими (рис. 35, Б). Интенсивная деятельность камбия, сопровождающаяся разрастанием сердцевины за счет растяжения ее клеток, приводит к очень быстрому утолщению стебля. В течение двух-трех месяцев стебель утолщается почти в 10 раз.
В старых стеблях стенки клеток сердцевины и лучевой паренхимы, расположенной между ксилемными участками пучков, подвергаются одревеснению, протопласты клеток отмирают.
Задание.
1. Зарисовать схему строения стебля подсолнечника, отметив эпидермис с волосками, первичную кору, состоящую из колленхимы и паренхимы, центральный цилиндр с кольцом открытых проводящих пучков и межпучковым камбием. Обратить внимание на разное строение проводящих пучков и на присутствие в некоторых из них протофлоэмных волокон. Отметить расположение схизогенных вместилищ выделений.
2. При большом увеличении зарисовать схизогенное вместилище выделений.
Стебель сныти обыкновенной (Aegopodium podagraria L.)
Для изучения наиболее пригодны участки стеблей с длинными междоузлиями, расположенными под нижними боковыми цветоносными веточками. Фиксировать материал лучше в конце июня —начале июля, когда стебли достаточно эластичны и не ломаются при изгибе.
Поперечные срезы делают с разрезанных вдоль кусочков стебля, обрабатывают их флороглюцином и соляной кислотой и рассматривают при малом и большом увеличениях микроскопа. На некоторых срезах желательно провести реакцию с раствором иода в водном растворе иодистого калия.
Стебель сныти полый, в поперечном сечении более или менее округлый, с небольшими тупыми выступами, наличие которых объясняется продольной ребристостью стебля.
Эпидермис типичного для этой ткани строения, с неровной блестящей желтоватой кутикулой. Некоторые’ его клетки образуют короткие одно-трехклеточные волоски. Под эпидермисом находятся один-два слоя клеток хлорофиллоносной паренхимы, к которой примыкают тяжи уголковой колленхимы. Наиболее крупные тяжи колленхимы проходят вдоль ребер.
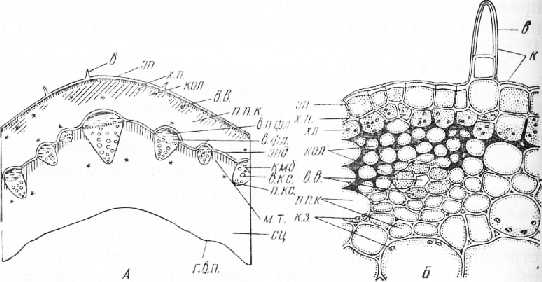
Пространство между участками этой механической ткани и под ними, до проводящих пучков, занимает паренхима, состоящая из крупных округлых клеток. В некоторых клетках видны крахмальные зерна. Больше всего их в самом внутреннем извилистом слое, дифференцированном в крахмалоносное влагалище. Хлорофиллоносная паренхима, колленхима и расположенная внутрь от нее крупноклеточная паренхима составляют первичную кору стебля.
Центральный цилиндр начинается кольцом открытых коллатеральных проводящих пучков. Проводящие пучки разных размеров. Наиболее крупные из них располагаются под выступами стебля. Наряду с полными пучками, содержащими флоэму и ксилему, нередко встречаются неполные, составленные только флоэмой, иногда в них развиваются один-два сосуда ксилемы.
Неодинаковые размеры проводящих пучков и разная степень развития в них проводящих тканей объясняются неодновременным заложением в образовательном кольце тяжей прокамбия, клетки которого в разное время начинают дифференцироваться в проводящие элементы. В ранее заложившихся прокамбиальных тяжах первичные проводящие ткани развиваются быстрее, в таких пучках раньше формируется камбий и начинается образование вторичных тканей.
Выбрав наиболее крупный проводящий пучок, следует подробнее ознакомиться с его строением при большом увеличении микроскопа. Наружная часть флоэмы представлена мелкими, плотно соединенными клетками с утолщенными, иногда блестящими стенками. В очертании клетки многоугольные. Они составляют наиболее старую по времени образования часть первичной флоэмы— протофлоэму. Стенки самых наружных клеток протофлоэмы могут одревесневать, и клетки превращаются таким образом в механические элементы. Чем старше стебель, тем сильнее армированы проводящие пучки.
Остальная часть флоэмы представлена ситовидными трубками с сопровождающими клетками и паренхимны-ми элементами. Ширина камбиальной зоны зависит от фазы развития растения: до цветения она обычно широкая, к началу цветения камбий постепенно прекращает деятельность, его производные дифференцируются в элементы флоэмы или ксилемы, камбиальная зона становится узкой. Если материал собран в конце вегетационного периода, камбиальная зона в пучках практически незаметна.
Большую часть ксилемы составляют элементы камбиального происхождения: крупнопросветные сосуды, толстостенные волокна (либриформ), паренхимные клетки. Все эти элементы имеют одревесневшие стенки и расположены плотно. Первичная ксилема представлена несколькими мелкими сосудами, окруженными живыми па-ренхимными клетками с тонкими целлюлозными оболочками. В некоторых пучках внутренняя часть протокси-лемы, граничащая с сердцевиной, содержит одревесневшие волокна, многоугольные в поперечном сечении. Кси-лемные части проводящих пучков соединены межпучковой одревесневшей механической тканью.
Сердцевина состоит из крупных тонкостенных паренхимных клеток. Наиболее мелкие клетки расположены вокруг пучков. В старых стеблях стенки клеток могут быть одревесневшими. При развитии стебля внут- 1 ренняя часть сердцевины разрушается и на ее месте воз- 1 никает крупная воздушная полость, которая проходит вдоль всего междоузлия.
В стебле сныти хорошо развита система вместилищ выделений схизогенного происхождения . Они встречаются в колленхиме и паренхиме первичной коры, во флоэме пучков и в сердцевине близ ксилемы. Вмести- { лище имеет вид межклетника, окруженного несколькими эпителиальными клетками с более густым содержимым, чем в соседних клетках.
Задание. Пои малом увеличении микроскопа зарисовать схему строения стебля, отметив эпидермис, хлоро- ; филлоносную паренхиму, участки уголковой колленхимы, крахмалоносное влагалище, окружающее кольцо открытых коллатеральных пучков, соединенных межпучковой механической тканью. Показать на схеме расположение ‘ вместилищ выделений.
- * *
Описанное строение стебля характерно для большинства представителей семейства зонтичных. Различия могут быть в степени развития колленхимы, ширине и степени одревеснения межпучковой ткани, числе и расположении вместилищ выделений.
Стебель лютика ползучего (Ranunculus repens L.)
Тонкий, полый внутри стебель лютика развивается в течение короткого времени, так как побег быстро переходит в репродуктивную фазу — фазу формирования цветков. Механические и проводящие ткани в нем развиты слабо. На поперечном срезе, обработанном флороглюцином и соляной кислотой, можно видеть, что проводящая система стебля представлена отдельными пучками, расположенными по кругу среди тонкостенных клеток паренхимы, окружающих крупную центральную полость, которая возникла вследствие разрушения вНутренней части сердцевины.
Эпидермис состоит из вытянутых по длине стебля, узких в поперечнике клеток со слегка утолщенными наружными стенками и слабо заметным слоем кутикулы на их поверхности.
Первичная кора сложена тонкостенными парен-химными клетками, содержащими хлоропласты. В коре имеются многочисленные межклетники. У живых растений они заполнены воздухом. Такую ткань называют аэренхимой. Она характерна для растений, развивающихся во влажных условиях.
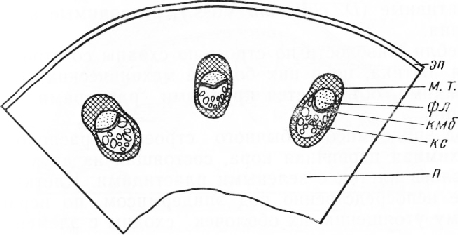
Резкой границы между первичной корой и центральным цилиндром нет, и паренхима первичной коры постепенно переходит в паренхиму центрального цйлиндра.
Проводящие пучки коллатеральные. В них хорошо различимы все составляющие их ткани. Во внутренней части пучка видны немногочисленные мелкие сосуды первичной ксилемы, окруженные тонкостенными паренхимными клетками. К периферии от них располагается вторичная ксилема, состоящая из более широко-просветных сосудов, небольшого числа механических волокон и паренхимных клеток с одревесневшими оболочками. Хорошо выраженная камбиальная зона из нескольких слоев тонкостенных клеток, расположенных радиальными рядами между ксилемой и флоэмой, заметна лишь в молодых стеблях. К началу цветения камбий обычно заканчивает деятельность и камбиальная зона становится незаметной. Тонкостенные элементы флоэмы представлены только ситовидными трубками с сопровождающими клетками. Пучки окружены одревесневшими толстостенными волокнами прокамбиально-го происхождения.
Задание. Зарисовать схему строения стебля, на которой следует обозначить расположение всех тканей.
С пучковым строением стеблей можно ознакомиться также на примере живокости (Delphinium), используя декоративные (D. cultorum Voss.), разводимые в садах растения.
Стебли живокости по строению сходны со строением стебля лютика, но в них больше механических тканей, что, видимо, объясняется крупными размерами этого растения.
Под эпидермисом обычного строения расположена паренхимная первичная кора, состоящая из двух, трех, пяти слоев клеток с зелеными пластидами. Клетки, лежащие непосредственно под эпидермисом, по неравномерному утолщению их оболочек сходны с элементами колленхимы.
В центральном цилиндре находится кольцо открытых коллатеральных пучков с тяжом одревесневших протофлоэмных волокон. Пучки соединены межпучковой одревесневшей паренхимой.
Во флоэме можно рассмотреть ситовидные трубки и сопровождающие клетки.
Вторичная ксилема представлена довольно крупными сосудами и небольшим числом волокон. Внутреннюю часть пучка составляют проводящие элементы первичной ксилемы, окруженные толстостенными клетками.
Камбиальная зона сложена таблитчатыми тонкостенными клетками, расположенными в три-пять слоев. Межпучковый камбий обычно заметен только в старых стеблях.
Стебель вьюнка полевого (Convolvulus arvensis L.)
На примере вьюнка можно ознакомиться с так называемым сплошным или непучковым типом строения центрального цилиндра стебля.
Такое строение проводящей системы возникает в том случае, если при развитии стебля прокамбиальные тяжи, дифференцирующиеся в образовательном кольце, разрастаясь, соединяются в полый цилиндр. Лишь в верхних, самых молодых междоузлиях стебля может сохраняться пучковое строение. Непучковое строение нередко называют кольцевым, однако этот термин нельзя считать удачным, так как «кольцом» проводящие ткани располагаются только на поперечных срезах.
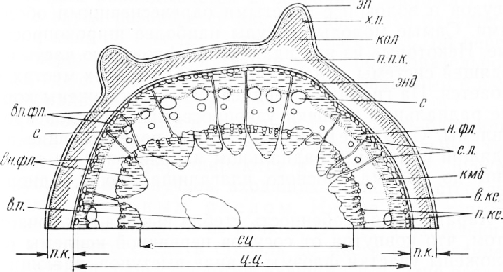
Стебель вьюнка в поперечном сечении округлый, с небольшими выступами (рис. 6). Снаружи он покрыт эпидермисом обычного строения. Глубже расположена очень узкая первичная кора, наружная часть которой представлена одним-двумя слоями тонкостенных паренхимных клеток с хлоропластами. Их подстилают два-три слоя клеток уголковой колленхимы. В ребрах колленхима более мощная. Под ней находятся тонкостенные паренхимные клетки. Первичная кора закапчивается крахмалоносным влагалищем.
В центральном цилиндре перицикла, видимо, нет. Отдельные группы волокон с неодревесневшими слабоутолщенными оболочками и паренхимными клетками между ними дифференцируются из периферической зоны прокамбия и должны быть отнесены поэтому к первичной флоэме. Флоэма, камбий и ксилема на поперечном срезе стебля расположены кольцами.
Флоэма состоит из ситовидных трубок с сопровождающими клетками и флоэмной паренхимы. Однако эти элементы настолько мелки, что их трудно различить.
Если материал собран в конце вегетационного периода, то камбиальная зона в стебле обычно незаметна, так как к этому времени камбий заканчивает деятельность.
Вторичная ксилема состоит главным образом из сосудов и волокон с толстыми одревесневшими оболочками. Самые молодые сосуды наиболее широкопросвет-ные. Некоторые из них вдаются во флоэмную часть проводящей системы, и камбиальная зона в этих местах изгибается. На границе с сердцевиной расположены короткие радиальные цепочки узкопросветных сосудов прото-ксилемы. Сердцевинные лучи, пересекающие проводящий цилиндр в радиальном направлении от сердцевины до крахмалоносного влагалища, узкие, однорядные.
Характерная особенность стебля вьюнка заключается в том, что ковнутри от сосудов первичной ксилемы расположены участки флоэмы в виде выступов, вдающихся в паренхимную сердцевину. По происхождению эта флоэма первичная, так как она образовалась из внутренних зон прокамбиальных тяжей в процессе их дифференцировки.
При рассмотрении стебля нужно обратить внимание на сравнительно слабое развитие механических элементов. Это связано с тем, что стебель вьюнка в качестве опоры использует другие растения, а при отсутствии опоры стелется по поверхности почвы.
Задание.
1. Зарисовать схему строения стебля, отметив особенности расположения составляющих его тканей.
2. При большом увеличении микроскопа зарисовать группу волокон в наружной флоэме.
- * *
Вместо вьюнка с непучковым строением центрального цилиндра можно ознакомиться на примере барвинка (Vinca), ластовня (Antitoxicum), дербенника (Lythrum), подмаренника (Galium), калистегии (Calystegia) и других растений.
Анатомия растений - Стебель травянистых двудольных растений