Строение «корнеплодов»
Наряду с функциями поглощения и проведения корни выполняют функцию запаса питательных веществ, которые иногда накапливаются в очень больших количествах. Таковы корни некоторых двулетних двудольных растений (морковь, петрушка, репа, редька, свекла и др.), образующие корнеплоды.
В состав корнеплода кроме корня входит подсемя-дольное колено (гипокотиль), а также морфологически нижняя часть побега. У разных «корнеплодов» эти органы развиты и утолщены в разной степени. Так, у моркови, петрушки, некоторых сортов редьки и сахарной свеклы большая часть «корнеплода» образована главным корнем; у репы, столовых сортов свеклы и других—-основанием стебля и гипокотилем, а собственно корень представляет собой лишь самую нижнюю часть «корнеплода», несущую боковые корни.
Запасные вещества, отложенные в «корнеплоде» в первый год жизни растения, на следующий год расходуются на развитие цветоносных побегов, плодов и семян.
Питательные вещества накапливаются в живых паренхимных клетках с тонкими неодревесневшими стенками. Клетки богаты клеточным соком, поэтому корнеплоды всегда сочные.
Вещества могут откладываться: 1) преимущественно в древесинной (ксилемной) паренхиме (редька, репа и другие крестоцветные); 2) в древесинной и особенно в лубяной (флоэмной) паренхиме (морковь, петрушка и другие зонтичные) 3) в паренхиме, образованной деятельностью нескольких добавочных камбиев (свекла).
Для изучения особенностей вторичного утолщения можно использовать корни толщиной от 2 до 5 мм. Поперечные срезы последовательно обрабатывают раствором иода в водном растворе иодистого калия, флороглю-цином, соляной кислотой и рассматривают при малом и большом увеличениях микроскопа.
Корень моркови (Daucus sativus (Hoffm.) Roehl.)
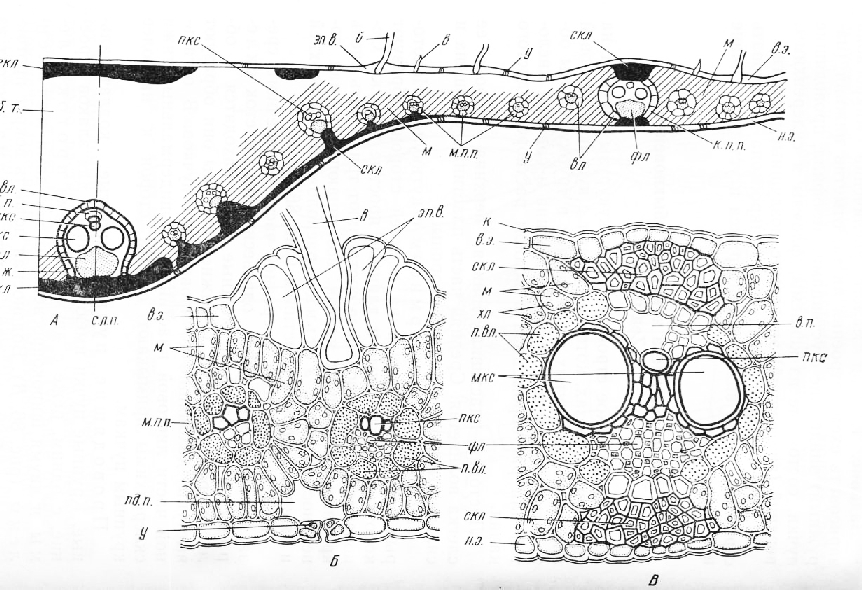
На поперечном срезе, еще не обработанном реактивами, хорошо различимы две зоны: внутренняя, довольно узкая, светло-желтая (вторичная ксилема) и наружная —более широкая, оранжевая (вторичная флоэма). Во флоэме сосредоточена основная масса крахмала, растворимых Сахаров и других питательных веществ . Оранжевый цвет обусловлен присутствием в ее клетках хромопластов с кристаллами каротина. Флоэма и ксилема разделены светлой камбиальной зоной, которая часто разрывается, и проводящие ткани отделяются одна от другой.
На срезе, обработанном флороглюцином и соляной кислотой, уже при малом увеличении микроскопа можно обнаружить два очень коротких, узких, обычно сходящихся в центре корня тяжа трахеальных элементов первичной ксилемы, окрасившихся в красный цвет.
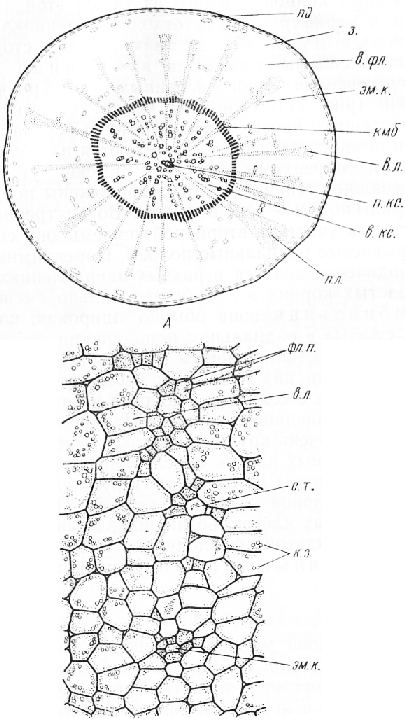
От наружных концов каждого из тяжей этой диархной ксилемы к периферии среза отходят крупноклеточные первичные лучи. Между ними, по обе стороны от первичной ксилемы, находится вторичная ксилема, пересеченная многочисленными вторичными лучами. Основная масса ксилемы состоит из тонкостенных паренхимных клеток. Сосудов в корне моркови немного. Они резко выделяются на общем фоне бесцветной паренхимы, так как имеют утолщенные одревесневшие стенки. Сосуды расположены поодиночке или собраны по три — семь в группы. Во внутренних, наиболее старых участках вторичной ксилемы они составляют прерывистые радиальные полосы. Паренхимные клетки периодически делятся в разных направлениях, поэтому в толстых корнях все элементы сильно смещены.
Камбиальная зона обычно широкая, клетки ее мелкие, сжатые в радиальном направлении.
Флоэма состоит главным образом из паренхимных элементов. Среди них находятся небольшие группы ситовидных трубок с сопровождающими клетками и схизо-генные эфирномасляные каналы. На срезе хорошо заметны лучи из очень крупных клеток. Периодические деления паренхимных клеток обусловливают значительное разрастание флоэмы в толщину. Наружная часть флоэмы граничит с зоной из нескольких (6—7) рядов крупных паренхимных клеток, возникших, вероятно, вследствие делений клеток феллодермы. В этой зоне видны эфирномасляные каналы. Корень покрыт тонким слоем пробки.
- * *
Корень моркови можно заменить корнем петрушки (Petroselinum sativum Hoffm.), имеющим сходное строение. Отличие состоит в том, что окружающая флоэму зона паренхимы с крупными эфирномасляными каналами у петрушки более широкая, хромопластов в клетках нет.
Корень редьки (Raphanus sativus L.)
В отличие от моркови утолщение корнй редьки происходит главным образом за счет сильного разрастания вторичной ксилемы. Вторичная флоэма по сравнению с мощной ксилемой развита весьма слабо.
В центре поперечного среза корня находятся мелкие сосуды диархной первичной ксилемы, составляющие короткую веретеновидную цепочку. От каждого ее конца, пересекая в радиальном направлении центральный цилиндр, отходит по одному широкому первичному паренхимному лучу.
Запасающая ткань корня представлена вторичной ксилемой, основная масса которой сложена тонкостенными клетками тяжевой и лучевой паренхимы; клетки ее содержат крахмал и богаты клеточным соком. Не многочисленные широкопросветные сосуды собраны но два — шесть в короткие радиальные цепочки, расширяющиеся к периферии. Вблизи камбия сосуды обычно окружены небольшим числом механических элементов со слабоутолщенными, иногда одревесневшими стенками.
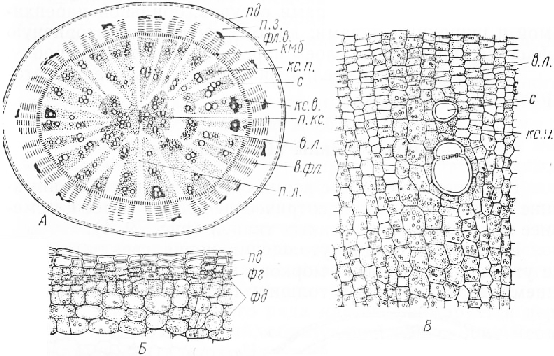
Камбиальная зона состоит из мелких таблитчатых клеток с густым цитоплазматическим содержимым. Снаружи она окружена узким кольцом вторичной флоэмы, в которой, как и в ксилеме, заметны широкие вторичные лучи. Вытянутые в радиальном направлении группы ситовидных трубок с сопровождающими клетками окружены паренхимой. К концу вегетации во вторичной флоэме выявляются тяжи коротких волокон. Их слабоутолщенные одревесневшие стенки имеют немногочисленные простые поры. Вокруг флоэмы расположен тонкий слой паренхимных клеток — производных перицикла и феллодермы. Корень покрыт бурой пробкой.
Задание.
1. Зарисовать схему строения корня, отметив диархную первичную ксилему; мощно развитую вторичную ксилему с группами сосудов, обильной паренхимой и широкими лучами; камбиальную зону; вторичную флоэму, окруженную слоем паренхимных клеток; пробку.
2. При большом увеличении зарисовать участок вторичной ксилемы.
Корень свеклы (Beta vulgaris L.)
На поперечном срезе корня обращают на себя внимание чередующиеся концентрические кольца более или менее интенсивно окрашенных тканей.
Начальные стадии утолщения корня свеклы сходны с утолщением корней моркови и редьки, но в дальнейшем его разрастание в толщину происходит своеобразно.
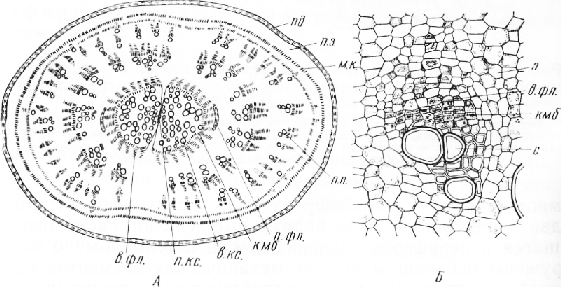
Корень проростка в фазе семядольных листьев имеет первичное строение. Радиальный проводящий пучок с диархной первичной ксилемой окружен однослойным перициклом. С появлением у растения первого настоящего листа начинает функционировать камбий, заложившийся между первичными ксилемой и флоэмой. В результате его деятельности на 12—15-й день жизни сеянца в центре корня по обеим сторонам от первичной ксилемы образуется по одному пучку, состоящему из вторичных ксилемы и флоэмы. Сильно обли-терированная к этому времени первичная флоэма оттесняется к периферии. Вторичная ксилема состоит из сосудов с одревесневшими стенками и небольшого числа паренхимных клеток. Между пучками вторичных тканей проходят широкие лучи. Деятельность этого первичного камбия вскоре заканчивается и дальнейшее утолщение корня продолжается за счет добавочных к а м б и е в, возникающих последовательно один за другим при непосредственном участии перицикла и его производных.
Паренхимные клетки перицикла, делясь преимущественно тангентальными перегородками, образуют многослойную меристематическую ткань, которая на поперечном срезе расположена кольцом. В периферической части этого кольца закладывается феллоген, клетки средней части дифференцируются в паренхимные элементы, а из внутреннего ряда клеток формируется первый добавочный камбий, образующий новую зону мери-стематических клеток. Наружный ряд клеток этой зоны в дальнейшем будет функционировать в качестве второго добавочного камбия. Внутренние клетки меристе-матической зоны, делясь, откладывают наружу тонкостенные клетки паренхимы и небольшие группы клеток вторичной флоэмы, внутрь — крупные клетки паренхимы и непосредственно под участками флоэмы — элементы ксилемы.
Таким образом, в результате деятельности первого добавочного камбия возникает широкое кольцо паренхим,ной ткани с погруженными в нее мелкими коллатеральными пучками. Флоэма состоит из нескольких ситовидных трубок с сопровождающими клетками и клеток паренхимы, ксилема —из небольшого числа пористых сосудов, окруженных механическими элементами, и ксилемной паренхимы.
Вскоре начинают делиться клетки второго добавочного камбия, образуя следующую меристематическую зону. Наружные клетки ее дифференцируются в третий добавочный камбий, а внутренние формируют второе кольцо проводящих пучков и межпучковую паренхиму. Третий добавочный камбий з свою очередь участвует в образовании четвертого добавочного камбия и третьего кольца проводящих пучков и т. д.
Деятельностью добавочных камбиев объясняется наличие на поперечном срезе корня нескольких (8 и более) концентрических колец, состоящих из мелких проводящих пучков и обильной паренхимы, в тонкостенных клетках которой накапливаются сахароза и другие питательные вещества. У сюловых сортов свеклы в паренхимных клетках много антоциана. Чем более удалены добавочные камбии от центра корня, тем слабее их деятельность. Поэтому ширина концентрических колец, число и размеры проводящих пучков уменьшаются к периферии среза. Самые молодые наружные кольца могут быть представлены лишь узкими слоями слабо-дифференцированных или даже меристематических клеток.
Интенсивность вторичного утолщения корня свеклы тесно связана с развитием листового аппарата. Установлено, что число концентрических слоев зависит от числа листьев в прикорневой розетке.
Деятельность добавочных камбиев заканчивается рано, и дальнейшее утолщение корня происходит за счет деления и разрастания паренхимных клеток.
Снаружи корня находится тонкий слой мелкоклеточной паренхимной ткани, окруженной темно-бурой пробкой.
Задание.
1. При малом увеличении микроскопа зарисовать схему строения корня, отметив на ней ди-архную первичную ксилему, первичные лучи, вторичные флоэму и ксилему, образованные деятельностью обычного камбия, и концентрические кольца паренхимной ткани с погруженными в нее мелкими проводящими пучками, отложенные добавочными камбиями, недифференцированные меристематические клетки периферических колец и пробку.
2. При большом увеличении микроскопа зарисовать участок паренхимной ткани с проводящим пучком, образованным добавочным камбием. лист
Типичный лист представляет собой пластинчатый орган растения, имеющий дорзовентральное строение: в нем различают верхнюю (брюшную) и нижнюю (спинную) стороны. Обычно лист состоит из пластинки и черешка.
Лист закладывается на верхушке побега под конусом нарастания в виде бокового выроста, из верхней части которого впоследствии развивается пластинка, а из нижней — основание листа. Затем путем интеркалярного (вставочного) роста между основанием и пластинкой развивается черешок.
В листе совершается процесс фотосинтеза, т. е. превращение неорганических веществ (углекислого газа и воды) в органические при обязательном участии световой энергии. В природе источником энергии служат солнечные лучи. Продукты фотосинтеза — углеводы — перемещаются из листа в другие органы растения. Лист осуществляет также испарение воды—транспирацию и газообмен с окружающей средой.
В соответствии с выполняемыми функциями в листе представлены следующие ткани: ассимиляционная, в клетках которой происходит процесс фотосинтеза, п о-кровная, регулирующая испарение воды и газообмен, проводящие и механические. Нередко встречаются идиобласты (одиночные клетки или группы клеток), механические или участвующие в отложении минеральных соединений или специфических органических веществ: эфирных масел, дубильных веществ, смол, каучука и т. д.
Строение листовой пластинки фикуса каучуконосного (Ficus elastica Roxb.)
Для ознакомления со строением листа весьма удобны листья фикуса — вечнозеленого растения, широко распространенного в комнатной культуре. С его толстых кожистых листьев нетрудно получить хорошие срезы. Из листовой пластинки сначала вырезают параллельно боковым жилкам полоску 0,5—1 см шириной, затем ее складывают 3—4 раза поперек и делают срезы перпендикулярно боковым жилкам. Срезы кладут на предметное стекло в каплю воды. При малом увеличении микроскопа косые и толстые срезы удаляют, остальные накрывают покровным стеклом.
С обеих сторон лист покрыт эпидермисом. Между верхним и нижним эпидермисом находится ассимиляционная ткань — мезофилл, клетки которого содержат многочисленные хлоропласта, обусловливающие зеленую окраску листа. В мезофилле расположены проводящие пучки с сопровождающей их механической тканью (рис. 4).
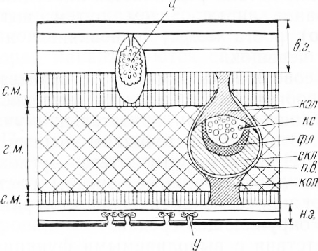
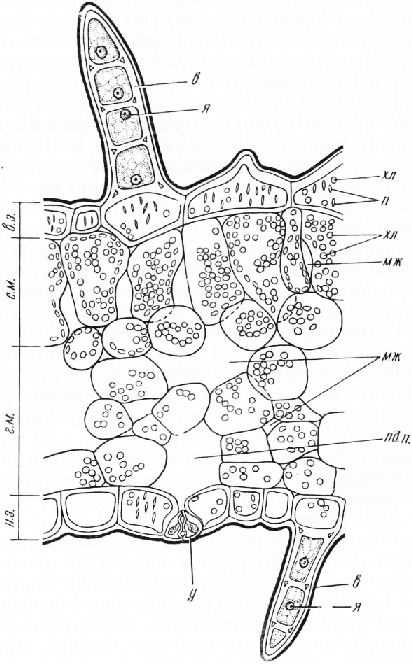
При большом увеличении микроскопа на верхней и нижней сторонах листа видны три слоя прозрачных клеток. Самый внешний слой состоит из мелких клеток с утолщенными наружными стенками, на поверхности которых хорошо заметна кутикула. Такое строение типично для хожицы, или эпидермиса. На нижней стороне листа в эпидермисе видны устьица, на верхней стороне их нет. Клетки второго и третьего слоев крупнее наружных клеток. Они имеют тонкие пористые стенки. Эти слоя клеток, которые обычно принимают за гиподермувозникают на ранней стадии развития листовой пластинки из клеток покровного слоя вследствие их делений тангентальными перегородками.
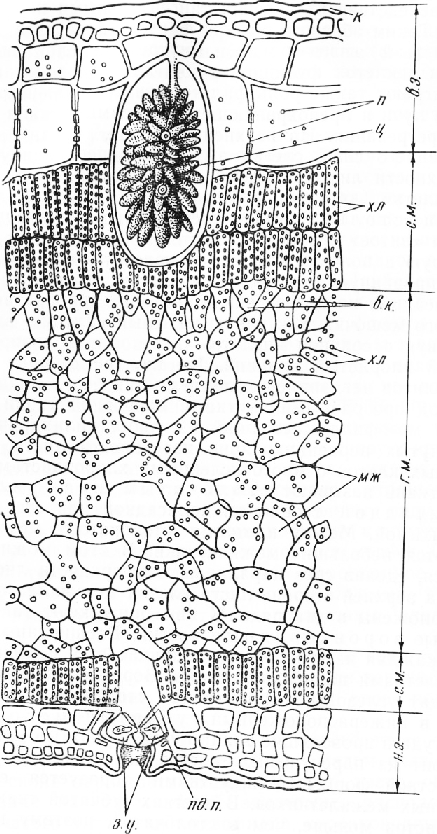
Таким образом, в листе фикуса образуется трехслойный эпидермис. Внутренние слои эпидер-мальных клеток функционируют как водозапасающие. Они играют также роль фильтра, задерживающего тепловые лучи и предохраняющего тем самым ткани листа от перегревания. На верхней стороне, среди клеток внутреннего слоя, слегка вытянутых перпендикулярно поверхности листа, нередко встречаются очень крупные клетки (литоцисты) с гроздевидными образованиями-— цистолитами (от греч. «китис» — пузырь, мешок и «литое» — камень). Цистолит представляет собой вросшую в полость клетки целлюлозную оболочку, инкрустированную углекислой известью. После проведения реакции на одревеснение цистолит приобретает вид смятого мешочка, так как углекислый кальций, взаимодействуя с соляной кислотой, переходит в водорастворимый хлористый кальций. На нижней стороне листа цистолитов нет, эпидермальные клетки более мелкие.
Под прозрачными клетками покровной ткани находится мезофилл, состоящий из узких цилиндрических, вытянутых перпендикулярно поверхности листа тонкостенных клеток с многочисленными хлоропластами. Такую ткань называют столбчатым мезофиллом, палисадной тканью, палисадной или столбчатой паренхимой. Между клетками столбчатого мезофилла имеются небольшие межклетники. В этом можно убедиться, сделав срез параллельно поверхности листа.
На верхней стороне листа клетки мезофилла обычно расположены в два ряда. К ним примыкают так называемые воронковидные клетки. Расширенным концом каждая из этих клегок соприкасается с несколькими клетками палисадной ткани. Воронковидные клетки отводят углеводы, выработанные столбчатым мезофиллом, в нижерасположенный губчатый мезофилл, а оттуда в проводящие ткани пучка. Губчатый мезофилл состоит из паренхимных клеток неправильной, часто лопастной формы. Между ними образуется система крупных межклетников. В клетках губчатой ткани хло-ропластов меньше, чем в столбчатой, поэтому нижняя сторона листа светлее верхней. Под губчатым мезофиллом находится столбчатый мезофилл из одного ряда клеток более коротких, чем клетки на верхней стороне листа.
В листовой пластинке хорошо развита сеть закрытых коллатеральных проводящих пучков. Лишь в средней жилке многолетних листьев иногда встречается камбий.
Ксилема, состоящая в основном из элементов со спиральными утолщениями, в листе всегда обращена к морфологически верхней стороне. Входя в стебель, пучок дуговидно изгибается, поэтому в стебле ксилема занимает внутреннее, а флоэма — наружное положение. Ориентация проводящих тканей в пучке — надежный критерий для определения морфологически верхней и нижней сторон листа. Пучки разных порядков ветвления имеют разные размеры и строение. Наиболее крупные пучки, представляющие собой ответвления от средней жилки, состоят из хорошо развитых флоэмы и ксилемы. Флоэму огибает тяж склеренхимы в виде желоба, который на вертикальном срезе имеет подковообразные очертания. Склеренхима состоит из волокон с толстыми одревесневшими стенками. Пучки окружены влагалищем, или обкладкой, из бесцветных тонкостенных паренхимных клеток. Сверху и снизу к крупным проводящим пучкам примыкают группы клеток с не-одревесневшими, часто неравномерно утолщенными стенками. Эти клетки, сходные по внешнему виду с элементами колленхимы, вплотную подходят к эпидермису. По мере разветвления пучки становятся мельче. Сначала они теряют склеренхиму, затем паренхимное влагалище, флоэму и заканчиваются отдельными трахеи-дами. Мелкие пучки могут быть разрезаны вдоль или косо.
Задание.
1. Со среза, заключенного в воду, зарисовать при большом увеличении микроскопа верхний трехслойный эпидермис. Во внутреннем слое показать клетку с цистолитом.
2. После проведения реакции на одревеснение с фло-роглюцином и соляной кислотой при малом увеличении микроскопа зарисовать схему строения листа, обратив внимание на трехслойный эпидермис, расположение столбчатого и губчатого мезофилла, воронковидных клеток; в нижнем эпидермисе отметить устьица; показать закрытый коллатеральный проводящий пучок со склеренхимой, огибающей пучок со стороны флоэмы, парен-химным влагалищем и колленхимоподобпой тканью, расположенной между пучком и эпидермисом.
3. При большом увеличении микроскопа зарисовать столбчатый мезофилл, воронковидные клетки и губчатый мезофилл.
Вместо фикуса можно использовать лист камелии японской (Camellia japonica L.). В листе камелии эпидермис однослойный, столбчатый мезофилл развит только на верхней стороне, некоторые клетки губчатого мезофилла содержат друзы оксалата кальция. В мезофилле встречаются крупные одревесневшие склереиды разной формы.
- * *
Дифференциация мезофилла на столбчатый и губчатый наблюдается в листьях растений, развивающихся в хороших условиях освещения. У растений тенистых местообитаний столбчатого мезофилла в листьях нет или он слабо развит. Для теневой структуры листа характерен мезофилл губчатого типа. Эпидермис состоит из крупных клеток, которые часто имеют хлоропласты. Кутикула обычно очень тонкая. Механические ткани развиты слабо.
Листья такого строения имеют копытень европейский (Asarum europaeum L., рис. 82), кислица обыкновенная (Oxalis acetosella L.), адокса мускусная (Adoxa moschatellina L.), широко распространенные в лесах средней полосы СССР.
Строение листа касатика германского (Iris germanica L.)
Мечевидный лист касатика (ириса) сложен вдоль средней жилки так, что морфологически верхняя сторона его обращена внутрь, а нижняя — наружу. Наверху края листовой пластинки срастаются, на остальном протяжении они свободны.
Поперечный срез обрабатывают раствором иода в водном растворе иодистого калия и при малом увеличении микроскопа прежде всего изучают анатомическую топографию листа.
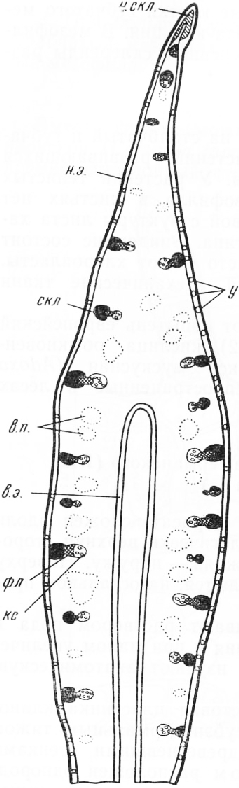
В средней части среза листовая пластинка клиновидно вытянута и снабжена субэпидермальным тяжом механических элементов с неодревесневшими стенками. Под эпидермисом расположен однородный мезофилл, состоящий из округлых тонкостенных паренхимных клеток, между которыми находятся многочисленные межклетники. Близ наружного (нижнего) эпидермиса клетки мезофилла мельче и богаче хлоропластами, чем клетки, расположенные на внутренней стороне листа. В толщу мезофилла погружены закрытые коллатеральные проводящие
пучки. Флоэма соприкасается с тяжом склеренхимы. В нижней части листа между пучками находятся крупные воздушные полости,
Ознакомившись с расположением тканей, следует при большом увеличении микроскопа подробно рассмотреть строение эпидермиса и устьичного аппарата.
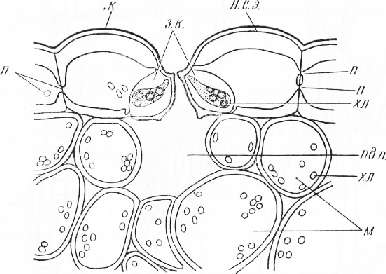
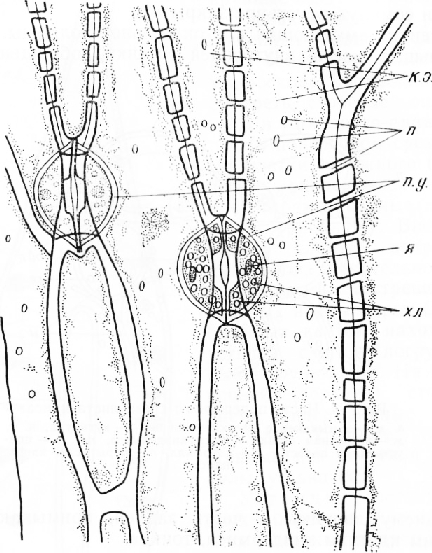
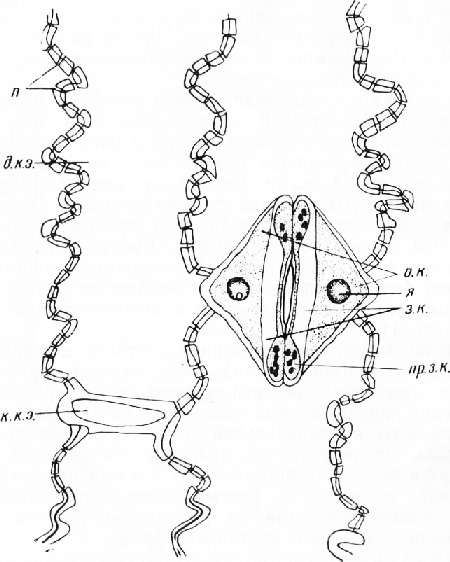
На поперечном срезе листа клетки наружного (нижнего) эпидермиса имеют почти прямоугольные очертания. Внешние стенки клеток, покрытые кутикулой, значительно толще боковых и внутренних, в которых видны поры. Среди обычных клеток находятся устьица, слегка погруженные в глубь листа. На внутренней (верхней) стороне листа эпидермальные клетки более крупные, тонкостенные, без заметной кутикулы; устьиц нет.
Устьице состоит из двух овальных замыкающих клеток с хлоропласта-ми. Между клетками находится межклетник — усть-ичная щель. Ее расширенную наружную часть называют передним двориком, а такую же расширенную внутреннюю — задним двориком. Он открывается в подустьич-ную полость. Внутренние стенки замыкающих клеток покрыты кутикулой. На поперечном срезе обычно хорошо заметны две пары кутикулярных выростов — клювиков. Одна из них ограничивает снаружи передний дворик, другая расположена у входа в подустьич-ную полость. На материале, фиксированном спиртом, устьичная щель почти не видна, так как замыкающие клетки сомкнуты (устьице закрыто).
Стенки замыкающих клеток сильно утолщены. Лишь небольшая часть внутренней стенки, обращенная к переднему дворику, и часть задней, примыкающей к другим клеткам эпидермиса, тонкие.
Строение эпидермиса и устьица можно рассмотреть также в плане. Для этого кожицу надрезают бритвой, осторожно сдирают ее с наружной стороны листа, кладут на предметное стекло в каплю воды или глицерина и накрывают покровным стеклом.
Клетки эпидермиса вытянуты по длине листа. Их боковые стенки несут многочисленные простые поры. Устьица расположены продольными рядами. Замыкающие клетки имеют очертания семени фасоли. Вогнутые, внутренние станки, обращенные к устьичной щели, сильно утолщены, наружные, выпуклые стенки — тонкие. В замыкающих клетках обычно хорошо видны хлоропласт ы.
Устьица регулируют испарение волы (транспирацию) и газообмен с внешней средой.
При избытке воды г. растении замыкающие клетки находятся в состоянии полного насыщения водой и имеют максимальный объем. Тонкие наружные стенки клеток сильно растянуты, клетки в середине несколько изогнуты, устьичная щель широкая (устьице открыто). При недостатке воды объем замыкающих клеток уменьшается, наружные стенки сокращаются, клетки выпрямляются, сближаются, устьичная щель становится уже (устьице закрывается).
Задание.
1. При малом увеличении микроскопа зарисовать схему строения листа, отметив нижний (наружный) эпидермис с устьицами, мезофилл, проводящие пучки, состоящие из ксилемы, флоэмы и склеренхимы, верхний (внутренний) эпидермис, воздушные полости.
2. При большом увеличении микроскопа зарисовать строение клеток нижнего эпидермиса и устьице в плане и в разрезе.
Строение листа кукурузы обыкновенной (Zea mays L.)
Анатомическое строение листа кукурузы изучают на поперечных срезах, которые делают с кусочков листовой пластинки, свернутых вдоль жилок в трубку. Срезы обрабатывают раствором флороглюцина и соляной кислотой или раствором иода в водном растворе иодистого калия.
Листовая пластинка тонкая, верхняя сторона ее слегка волнистая. Средняя часть пластинки утолщена и сильно выдается с нижней стороны.
Лист покрыт эпидермисом с кутикулой. Некоторые клетки эпидермиса на верхней стороне образуют короткие, шиловидные и длинные простые волоски. Основания длинных волосков составляют три — пять очень крупных прозрачных клеток, возвышающихся над поверхностью листа. Устьица расположены на верхней и нижней сторонах пластинки.
Под эпидермисом, в средней части поперечного среза, расположены тяжи одревесневших клеток склеренхимы. Наиболее мощные тяжи находятся в области средней жилки. С нижней стороны они образуют выступы, доходящие до проводящих пучков. В остальной части листовой пластинки субэпидермальные тяжи склеренхимы примыкают с двух сторон к наиболее крупным пучкам.
Проводящие пучки закрытые, коллатеральные. Крупные пучки имеют типичное для злаков строение, с одним, реже с двумя сосудами протоксилемы и двумя широкопросветными сосудами метаксилемы. Водоносная полость вокруг протоксилемы небольшая или ее нет. Флоэма состоит из ситовидных трубок, чередующихся с мелкими ,богатыми цитоплазмой сопровождающими клетками. Каждый пучок окружен влагалищем из округлых тонкостенных клеток. Паренхимные клетки влагалища, расположенные вокруг наиболее крупных пучков, в старых листьях нередко частично или полностью одревесневают.
В мелких проводящих пучках ксилема состоит из двух-трех одревесневших элементов, многоугольных в поперечном сечении. Иногда развита только флоэма. Паренхимное влагалище хорошо выражено, его клетки содержат хлоропласты.
Мезофилл однородный, состоящий из паренхимных клеток. На верхней стороне листовой пластинки клетки соединены довольно плотно и расположены венцом вокруг проводящих пучков. Близ нижней стороны листа между клетками мезофилла имеются крупные межклетники. На верхней стороне листовой пластинки, в средней утолщенной части, где проходит наиболее крупный пучок, мезофилла нет. Вместо него развита бесцветная ткань из крупных прозрачных тонкостенных клеток, бедных содержимым. Проводящие пучки, окруженные мезофиллом, смещены к нижней стороне листа.
Ознакомившись со строением листовой пластинки на вертикальном (поперечном) срезе, следует сделать поверхностный срез с нижней стороны листа и исследовать строение эпидермальных клеток в плане. Можно попытаться снять препаровальной иглой тонкий кусочек эпидермиса, предварительно надрезав его бритвой. Срез кладут в воду. Для просзетления его можно обработать соляной кислотой и рассматривать в глицерине, при этом желательно выбирать прозрачные участки среза, на которых не видны клетки мезофилла.
Эпидермис состоит из двух типов покровных клеток: длинных, вытянутых вдоль листовой пластинки, с сильно извилистыми стенками, и коротких — мелких, контуры которых почти прямоугольные. Короткие клет-
Устьица располагаются продольными рядами.
Замыкающие клетки в плане имеют вид узких прямоугольников с закругленными концами. Стенки в средней части каждой из замыкающих клеток сильно утолщены. Полость клетки в этом месте превращена в узкий канал, соединяющий тонкостенные концы клетки. Принцип работы устьичного аппарата злаков такой же, как и устьиц обычного строения. При насыщении водой форма замыкающих клеток изменяется, их концевые участки увеличиваются в объеме, вздуваются, устьичная щель расширяется. Потеря воды приводит к уменьшению объема тонкостенных участков замыкающих клеток, клетки восстанавливают прежнюю форму, устьице закрывается. К каждой замыкающей клетке сбоку примыкает по одной крупной околоустьичной клетке, которая в плане имеет треугольные очертания.
Задание.
1. При большом увеличении микроскопа зарисовать участок листа, показав эпидермис с волосками на верхней стороне пластинки и проводящий пучок с паренхимным влагалищем.
2. При большом увеличении зарисовать эпидермис в плане, отметив длинные клетки с извилистыми стенками, короткие клетки, а также устьичный аппарат, состоящий из замыкающих и околоустьичных клеток.
Строение листа ковыля перистого (Stipa pennata L.)
Растения, развивающиеся в условиях недостаточного водоснабжения, называют ксерофитами. Многие из них имеют жесткие мелкие листья. Ксероморфная структура характерна для листьев ковыля, растущего в степях, полупустынях и пустынях.
Для получения поперечных срезов фиксированные спиртом листья зажимают в сердцевину бузины или режут сразу пучок из 10—12 листьев. Срезы обрабатывают флороглюцином, соляной кислотой и заключают в глицерин.
Верхняя сторона листовой пластинки ребристая. Крупные ребра в поперечном сечении почти квадратные; чередующиеся с ними более мелкие ребра в очертании треугольные или округлотреугольные. Нижняя сторона листа ровная.
Лист покрыт эпидермисом с кутикулой. Стенки клеток обычно одревесневают. Некоторые клетки образуют короткие волоски в виде шипиков. Наиболее крупные шипообразные выросты находятся по краям листовой пластинки.
На дне ложбинок, расположенных между ребрами, три — пять эпидермальных клеток очень крупных, прозрачных. Расширенные концы этих клеток слегка погружены в мезофилл, суженные — обращены наружу. Боковые и внутренние стенки клеток тонкие, внешние — утолщены и покрыты кутикулой. Тяжи этих клеток, называемых пузыревидными, проходят вдоль листовой пластинки. На боковых сторонах ребер, возле их основания, расположены многочисленные мелкие устьица, защищенные волосками от действия солнечных лучей.
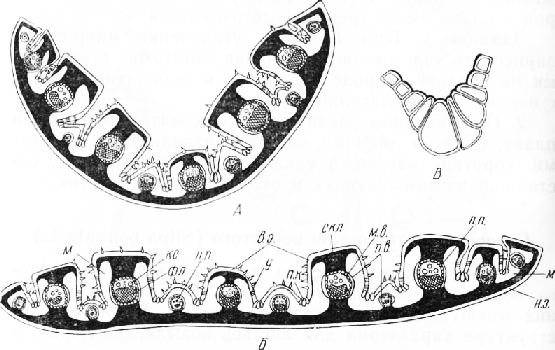
К нижнему эпидермису примыкает слой склеренхимы, состоящий из двух-трех рядов толстостенных одревесневших волокон. Внутренняя граница этого слоя неровная из-за выступов, которые входят в ребра, достигая флоэмной части находящихся в них пучков. Суб-эпидермальные тяжи склеренхимы, расположенные на верхних, плоских сторонах крупных ребер, также образуют выступы, соприкасающиеся с проводящим пучком со стороны ксилемы. На поперечном срезе склеренхим-ные тяжи, пересекающие крупные ребра, имеют вид двутавровых балок с крупным пучком посередине.
Проводящие пучки закрытые, коллатеральные, типичного для злаков строения. Пучок окружен двумя влагалищами: внутренним, местомным, состоящим из толстостенных одревесневших клеток, плотно прилегающих к проводящим тканям, и наружным, па-ренхимным, ограничивающим пучок с боков. В мелких ребрах проводящие пучки слабо дифференцированы, ксилема в них развита плохо, иногда ее нет. Склерен-химный тяж примыкает к пучку лишь с нижней стороны.
Вся остальная часть листа, свободная от механической ткани, занята мезофиллом, состоящим из однородных, плотно сомкнутых хлорофиллоносных паренхимных клеток. На поперечных срезах зона, занятая мезофиллом, очертаниями напоминает латинские буквы V или W.
При недостатке воды листовая пластинка может свертываться в трубку. По мнению одних исследователей, гигроскопические движения объясняются падением тур гор а и уменьшением объема всех тонкостенных клеток, в том числе и пузыревидных клеток эпидермиса. Другие исследователи главную роль отводят сокращению склеренхимных тяжей, происходящему при обезвоживании стенок волокон. Возможно, что пузыревидные клетки несколько облегчают свертывание листа.
Задание.
1. При малом увеличении микроскопа зарисовать схему строения листа, отметив на ней эпидермис с устьицами, волосками и пузыревидными клетками, склеренхиму, проводящие пучки с двумя влагалищами, мезофилл.
2. При большом увеличении микроскопа зарисовать строение пузыревидных клеток.
Анатомия растений - Строение «корнеплодов»