Шведская муха
В Европе, Азии и Северной Америке повреждаются шведской мухой кормовые травы, озимые хлеба, но особенно сильно — яровые; были нередко случаи, что посевы ячменя и яровой пшеницы полностью погибали от шведской мухи или давали очень низкий урожай.
Шведская муха (Oscinosoma frit L.), синонимы Osci-nis, Oscinella, относится к семейству Chloropidae— злаковые мушки (отряд двукрылые).
Шведская муха глянцевито-черного цвета, размер 1,5—2 мм. Имеет небольшой хоботок с широкими сосальцами, усики черные, на округлом третьем членике щетинка простая. Среднеспинка слегка выпуклая, без бороздок или продольных полос. Щиток сзади закругленный, крылья прозрачные, с металлическим отблеском, краевая жилка (костальная) доходит до четвертой продольной (медиальной). Ноги черные; брюшко черное, снизу желтоватое, у самки более толстое, заканчивающееся узким яйцекладом.
О. pusilla Meig. — передние и средние голени светлые, задние посредине затемнены. Более распространена на ячмене, пшенице, кукурузе и названа поэтому ячменной мухой (С. Н. Селиванова, Э. П. Нарчук). О. frit преобладает на овсе и названа овсяной мухой.
Шведская муха расселилась там, где произрастают кормовые растения — культурные и дикие злаки. На севере ее распространение и размножение ограничивает низкая температура, на юге — исключительная сухосты и жара. Следовательно, для размножения шведской мухи необходимы наличие оптимальной температуры, влажности и достаточное количество корма. При благоприятном сочетании этих трех факторов она размножается в массовом количестве. Однако решающим условием ее жизни, за исключением Крайнего Севера, является влажность.
В местности, где количество осадков за год меньше 200 мм, шведская муха не размножается или находится в минимуме; там же, где осадков больше 500 мм, шведская муха находит оптимальные условия для своего развития. Например, в степях Поволжья (Красный Кут, Уральск) осадков выпадает 150—250 мм, и шведская муха в этих местах вредит незначительно, тогда как в северо-западной части Саратовской области (Петровск, Сердобск, Аткарск) при наличии 450 мм осадков является весьма опасным вредителем злаков (Н. Л. Сахаров).
На территории Советского Союза шведская муха встречается в неодинаковой численности. Установлены следующие зоны вредности.
I. Зона наибольшего и постоянного вреда охватывает Белорусскую ССР и центральные области нечерноземной полосы (Московская, Владимирская, Тульская, Рязанская, Ивановская, Горьковская, Калининская, Калужская, Ярославская, Смоленская и Брянская). Эта зона смешанных, хвойно-широколиственных лесов. Здесь повреждаемость стеблей яровых хлебов доходит до 40—50% (А. И. Карпова, П. Г. Чесноков, И. М. Беляев, Е. Н. Киселева и др.).
II. Зона неустойчивого вреда характерна тем, что максимально шведская муха размножается лишь в отдельные, наиболее благоприятные годы; в северной части зоны имеют решающее значение температурные условия, а в южной — влажность. В эту зону входят северные области нечерноземной полосы (Ленинградская, Вологодская, Кировская, а на юге — степные районы Орловской, Курской и Воронежской областей, Украины и Поволжья).
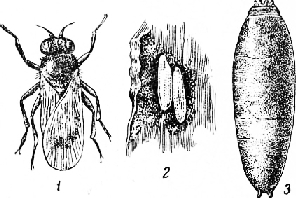
III. Зона слабого вреда: на севере — Карельская АССР, Коми АССР, а на юге — Ростовская, Волгоградская, Астраханская области. В данной зоне еще с большей резкостью выделяются на севере — холодные, а на юге — засушливые районы (степи и полынно-злаковые полупустыни).

Места обитания. Шведская муха выбирает места главным образом с низкой злаковой растительностью, хорошо освещенные и прогреваемые солнцем, защищенные от ветра. Такими излюбленными местами являются заросшие злаками дороги, где травяной покров не выше 5—10 см, дикие злаки вблизи полей (А. Н. Васина, 1929), луговые лужайки (А. Н. Швецова, 1930). В большом количестве мухи встречаются на скошенном лугу и даже на стогах сена.
Высокая численность ячменной мухи наблюдается на пастбищах, где процесс кущения злаков проходит в течение всего летнего периода (Э. П. Нарчук, 1955).
Кроме дикой, низкорослой растительности, шведская муха обитает на культурных злаках (яровые и озимые хлеба, кормовые травы). В течение вегетации злаков происходят миграции мух между культурными злаками и дикими в зависимости от состояния травостоя.
В начале роста овес, ячмень и другие хлебные злаки обыкновенно слабо заселяются шведской мухой (меньше, чем на меже). С наступлением кущения злаков численность мух на посевах заметно увеличивается до тех пор, пока не сомкнётся травостой и растения станут высокими. В период формирования трубки количество мух уменьшается и в период колошения достигает минимума, тогда как на меже в это время их чрезвычайно много. Когда зацветет овес, мухи снова появляются здесь и держатся в заметном количестве в течение всего периода цветения, после чего они переселяются на дикие злаки.
К временным местам обитания можно отнести те растения, куда в период цветения мухи прилетают кормиться. Хотя травостой этих растений может быть очень густой и высокий, мухи все же встречаются на них в заметном количестве. Например, на 200 взмахов сачком попадалось па вике в период цветения 18—23 мухи, а на гречихе от 40 до 166.
В расселении шведской мухи на посевах, помимо состояния растительности, может иметь значение направление склона участка. М. А. Илюхиным (1959) установлено, что на южных склонах хлебные злаки сильнее повреждаются, чем на северных; вылет мух начинается раньше на южных склонах.
Определение количества мух при кошении сачком показало, что колебания численности насекомых связаны с состоянием погоды. Это видно из данных таблицы 19. В таблице приведены дни, близко стоящие, но с резкими колебаниями по числу пойманных мух. Например, 21 июня выловлено на 200 взмахов сачком 4 шведские мухи, 26 июня —194, 29 июня —только 29, а 1 июля снова повышение—100 мух. В момент кошения учитывалась температура на поверхности почвы, сила ветра, облачность и относительная влажность воздуха. Снижение количества мушек в одних случаях зависит от понижения температуры, в других —от большой силы ветра; облачность также имеет значение.
В теплую, солнечную погоду мухи значительно подвижнее, легко вспугиваются и больше улавливаются сачком, тогда как в холодную погоду они укрываются и прочно удерживаются на растительности.
Количество личинок шведской мухи можно определить по числу поврежденных стеблей (обыкновенно в поврежденном стебле находится одна личинка).
Численность личинок меняется в зависимости от местоположения посева, сорта и срока посева (на раннем посеве 15 личинок, на позднем в 4 раза больше — 61 на 1 м2).
Перелеты шведской мухи. Для практических целей необходимо знать способность шведской мухи к перелету, дальности полета, ее расселение на посевах и т. д. Все эти вопросы мало изучены. На основании работ М. X. Шестерикова сложилось мнение, что шведская муха «летает плохо», бросками; далеких полетов не предпринимает. Такое утверждение достоверно, если наблюдать за шведской мухой утром или в холодную погоду.
В жаркую погоду можно наблюдать обратное: пойманные в сачок мухи очень подвижны, быстро из него улетают. Опыты Риддерта показали, что мухи летают на высоте 5—18 м, в зависимости от погоды; в холодную погоду отмечены на высоте около 18 м. Перелетам мушек способствует ветер, направляя и помогая им при полете.
Интересные опыты были проведены в США по определению расстояния перелетов мушки-дрозофилы (размер ее одинаков со шведской мухой).
Радиоактивные мухи были отмечены на расстоянии 7 км через 24 часа после выпуска. При легком преобладающем ветре мухи пролетали до 16 км в час.
Питание мух. Хоботок у шведской мухи небольшой, с широкими сосальцами, при помощи которых засасывается жидкая пища — нектар цветов. Для мух в качестве корма служит нектар многих видов растений. Нам приходилось отмечать мух на цветках гречихи, вики, люпина, сурепки, аниса и птичьей гречихи.
И. М. Замбии и А. И. Карпова (1940) отмечают, что шведскую муху наиболее привлекают в период цветения подмаренник, тысячелистник, одуванчик, василистник и валериана лекарственная.
По-видимому, кроме сахаристых веществ, находящихся в нектаре, шведская муха питается и белковой пищей. Неоднократно приходилось наблюдать при кошении сачком на ржи, овсе, мятлике значительное число мух в период цветения этих растений. Мы предположили, что пыльца злаков привлекает мушек как пища. Для проверки были помещены в садки с мушками яровая пшеница и тимофеевка в фазе цветения. Оказалось, что мухи, питавшиеся пыльцой, собираемой с пыльников, прожили 19—22 дня и откладывали яйца.
И. М. Замбии и А. И. Карпова также отмечают, что шведская муха питается белковой пищей, в частности пыльцой злаков — овса, ячменя и яровой пшеницы. Кроме того, И. М. Замбиным и И. Ф. Павловым установлено, что пищей для мух может служить «пасока»— сладкие выделения вегетативных частей злаковых растений.
Без пищи мухи могут жить очень непродолжительное время: в наших опытах вылетавшие из ложнококо-нов мухи могли прожить без пищи 2—3 дня, максимум 7, тогда как питавшиеся медом жили 20—47 дней, а получавшие корм с цветков сурепки — 13—28.
Размножение вида. Для определения зрелости мушек, их плодовитости и т. п. приходится прибегать к вскрытию самок, изучать состояние яичников. Это особенно необходимо для составления прогноза — начала появления шведской мухи, периода максимального лёта и откладки яиц, определения потенциальной плодовитости самок.
Нами изучалось созревание яичников у шведской мухи путем вскрытия самок на часовом стекле под лупой.
Рядом исследователей установлена прямая связь жирового тела с развитием яичников. При вскрытии брюшка шведской мухи на часовом стекле с небольшим количеством воды обыкновенно выходят дольки жирового тела в виде белых круглых капелек более или менее одинакового размера, которые легко можно подсчитать. Из ложнококонов молодые самки выходят с недоразвитыми яичниками, но с большим запасом жирового тела, в среднем 367 капелек. Яичники имеют вид двух маленьких клубочков с неразвитыми яйцами. В дальнейшем происходит заметное изменение и рост яиц, особенно тех, которые находятся ближе к яйцеводу. Яйцевые трубки содержат яйца различной зрелости. Соответственно развитию яиц постепенно убывает жировое тело; под конец жизни весь запас жирового тела бывает израсходован. По количеству жирового тела можно установить возраст самок. По нашему определению (1926— 1928 гг.), развитие яиц в яичниках проходит 5 стадий, и каждой стадии соответствует определенное количество жирового тела (число капелек), что можно видеть из данных таблицы 20,
Э. П. Нарчук (1955) при определении физиологического возраста по количеству жирового тела, наличию желтых тел в яичниках установила у ячменной шведской мушки также пять возрастных категорий. Вскрытие самок помогает точнее установить динамику развития шведской мухи в природе: начало лёта и интенсивность откладки яиц. Все эти моменты в жизни шведской мухи необходимо знать для составления прогноза и проведения оперативных мероприятий.
Молодые самки, имеющие больше жирового тела, более плодовиты. Нами установлено, что с увеличением жирового тела на 100 капелек число яиц в яйцевых трубочках повышается примерно на 3.
Спаривание происходит несколько дней спустя после вылета самок и зависит от зрелости яичников. В литературе отмечается, что спаривание может повторяться несколько раз.
По материалам вскрытия самок можно судить, что партия яиц, находящаяся ближе к яйцеводу, созревает не всегда одновременно: имеется несколько яиц (1—3), готовых к откладке, остальные 12—15 находятся на предшествующей (IV) стадии развития. В связи с этим яйца откладываются через определенные промежутки. Развитие яичников у шведской мухи длится не “менее 3—5 дней и зависит в значительной мере от температуры.
По нашим наблюдениям, непосредственно в природных условиях период от начала лёта мушек до начала яйцекладки продолжался, в зависимости от условий погоды, в одни годы 3—4 дня, в другие свыше 13 дней.
Место откладки яиц. На молодые стебли шведская муха откладывает яйца главным образом за колеопти-ле. Наблюдения и полевые опыты показали, что в оптимальную фазу для заражения (второй лист) максимальное количество яиц откладывалось за колеоптиле; очень немного отложено яиц на колеоптиле, на стебле, листьях и на земле вблизи растения.
Откладка яиц за колеоптиле имеет большое значение для выживаемости вида. Находясь под прикрытием колеоптиле, яйца гораздо лучше защищены от действия неблагоприятных внешних условий. Однако существует некоторая подвижность в выборе места откладки яйца, что связано всецело с состоянием колеоптиле,
По нашим наблюдениям, на всходах овса в фазе первого листа больше всего яиц (75%) откладывается на колеоптиле, поскольку оно еще плотно прилегает к стеблю и самке трудно провести за него свой яйцеклад. В фазе второго листа колеоптиле несколько отходит от стебля, и 83—93% яиц откладывается за него. Как только начинают появляться боковые стебли, яйца (19— 20%) откладываются за листовое влагалище, прикрывающее молодой стебель.
У ржи картина несколько иная: поскольку колеоптиле вначале неплотно прилегает к стеблю даже в фазе первого листа, яйца откладываются за колеоптиле, а далее, что очень характерно, в период кущения ржи большое количество яиц откладывается за листовое влагалище (47—65%). Степенью прижатости колеоптиле к стеблю можно объяснить выбор места откладки яйца в различные фазы развития растения.
По мере роста молодых растений колеоптиле все более отходит от стебля. В связи с этим мухи сначала откладывают больше яиц на колеоптиле, а затем почти все яйца откладывают за колеоптиле.
В наших опытах по избирательной способности самок выяснилось, что шведская муха предпочитает озимые культуры, так как по степени прижатости колеоптиле они более пригодны для откладки яиц.
На всходах овса в фазе второго листа больше всего встречалось по одному яйцу «а растении (68—69%), реже по два (21%) и только в отдельных случаях было отложено по 6—8 яиц, что, по-видимому, связано с кладкой не одной, а нескольких самок.
Прежде чем отложить яйца, самка тщательно исследует растение. Можно полагать, что шведская муха в основном при помощи осязания определяет пригодность стеблей для откладки яиц.
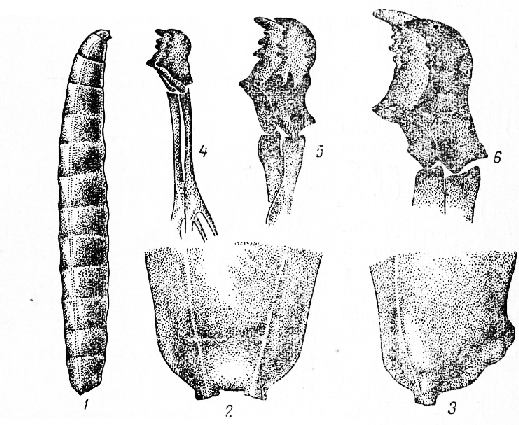
Развитие шведской мухи. Яйцо шведской мухи белое, продолговатое, около 0,7 мм длины и 0,2 мм ширины. Оболочка яйца покрыта продолговатыми бороздками, которые в средней части идут почти параллельно, а на концах постепенно сливаются; на притуплённом конце находится воронкообразное отверстие —микропиле, через которое проходит сперматозоид самца, оплодотворяющий яйцо. Рядом энтомологов установлена продолжительность развития яйца 3—8 дней, что в сильной степени связано с -состоянием окружающей температуры.
По данным Н. Н. Троицкого, в политермостате при температуре 18° развитие яйца проходит в течение 8—12 дней, при 22°—4—8 дней.
К концу развития яйца, как отмечает Н. В. Андреева, можно наблюдать через оболочку движения жвал сформировавшейся личинки, при помощи которых она прорывает возле микропиле яйцевую оболочку и выбирается наружу.
Вышедшей из яйца личинке не приходится долго искать корм, стоит лишь пробраться внутрь стебля, укрыться там от беспокоящего ее света, и она найдет у основания молодого стебля мягкие сочные ткани растения для своего питания.
Личинка -белая с желтоватым оттенком; форма тела цилиндрическая; передний конец тонкий, задний — округлый с двумя небольшими цилиндрическими выступами, открывающимися на вершине дыхальцами; на переднем конце тела, с нижней стороны третьего сегмента, расположены передние дыхальца, которые имеют вид веера из 5—6 дыхательных трубочек (число характерно для данного вида). Голова личинки не выделяется, усики короткие двучлениковые, ротовые крючки серповидные, зазубренные, у молодых личинок светло-коричневые, позднее становятся черными. Тело личинки состоит из
13 ясно разграниченных сегментов; взрослые личинки около 3—4 мм длины, 0,4 мм ширины. Личинки шведской мухи весьма сходны с другими личинками мух, обитающими также внутри стеблей злаков.
Молодая личинка шведской мухи, только что вышедшая из яйца, бесцветная и прозрачная, длиной около 1 мм. Через 4—6 дней личинка линяет. Продолжительность второго возраста 4—6 дней, размер личинки 1,5—3 мм.
В третьем возрасте (длится 10—16 дней) личинка становится желтой, ротовые крючки приобретают черную окраску, число зубцов увеличивается до 4—5, размер личинки 2,5—4,5 мм (Андреева и др.).
Жизнь личинки тесным образом связана с температурой. По наблюдениям Е. А. Крейтер, личинки неподвижны, если температура ниже 6°, слабое движение начинается при 7° и активное передвижение —при 10—12°.
Личинки начинают питаться, когда окружающая температура не ниже 12—14°.
В качестве корма личинки служат только сочные и нежные ткани, находящиеся в нижней этиолированной части стебля.
Исследованиями И. Д. Шапиро (1960) установлено, что личинки шведской мухи имеют крупные слюнные железы, которые выделяют фермент, растворяющий нежные растительные ткани. Этим осуществляется внеки-шечная подготовка пищи, в результате чего личинка оказывается среди разжиженного, высокопитательного пищевого субстрата.
Следует отметить, что количество и состав пищи, степень ее загрубения и влажность отражаются не только на росте и развитии самих личинок, но и на размере куколок и взрослых мушек, а затем на плодовитости самок.
Нам приходилось наблюдать, как из недовыкормив-шихся личинок развивались мелкие мушки, почти вдвое меньше нормальных. Однажды в сухую весну много стеблей у перезимовавшей озимой пшеницы высохло, личинки рано закуклились и впоследствии дали потомство мелких мушек.
Теперь становится ясной причина изменения количества яйцевых трубочек у самок и разносе количество жирового тела; здесь устанавливается прямая связь с питанием личинок.
Развитие личинки продолжается 18—26 дней; перед окукливанием личинка поворачивается головой вверх и становится неподвижной, при этом тело ее укорачивается (Н. В. Андреева).
Коконирование начинается при температуре 14° и происходит чаще всего внутри поврежденного стебля.
Личинка третьего возраста перед началом закукли-вания не сбрасывает свою шкурку, которая позднее затвердевает, приобретает желтую или коричневую окраску, в результате получается прочный ложнококон. Снаружи его видны личиночные органы: сзади выступы, а в передней части ротовые крючки и дыхальца. Длина ложнококона 2—3 мм, ширина 1,3 мм; у самок размер несколько больше, чем у самцов. Через 11—25 дней вполне сформировавшаяся мушка при помощи головного пузыря прорывает стенку ложнококона и выходит наружу; через час после вылета головной пузырь втягивается (М. К. Шестериков), крылья расправляются и затвердевают, мушка становится способной к полету.
Поколения шведской мухи. Типовой ход развития шведской мухи, по данным Н. Н. Троицкого (Пушкинская экспериментальная станция), проходит в следующем виде:
Весь цикл развития одного поколения на широте Ленинграда протекает 32—59 дней.
На основании исследований Пушкинской экспериментальной станции и Полтавской опытной станции можно считать, что развитие шведской мухи происходит при температуре не ниже 12—14°, действие эффективных температур начинается с этого уровня.
В течение летнего сезона развивается несколько поколений шведской мухи, при этом одно поколение накладывается на другое, отчетливой грани между ними нет, что очень затрудняет определение их числа.
Чтобы определить основные поколения шведской мухи, необходимо учитывать массовую откладку яиц в течение летнего периода. От ранних мушек, появившихся в третьей декаде мая (23—28) и отложивших яйца в начале июня, может развиться 3 поколения в год, но массовая откладка обыкновенно проходит во второй половине июня, следующие мушки из этих яиц появляются дней через 40, то есть в конце июля—-начале августа, а в середине —конце августа происходит вторая массовая откладка яиц; личинки, вышедшие из этих яиц, уходят на зимовку. В окрестностях Москвы развиваются два основных поколения: весеннее на яровых и осеннее на озимых и диких злаках; летнее поколение большей частью бывает в незначительном количестве.
В южной части СССР, где теплый период значительно дольше и сумма температур более высокая, число поколений шведской мухи увеличивается. В Воронежской области шведская муха дает 3 поколения в год (А. В. Жуковский, 1931).
Появление шведской мухи весной зависит от хостоя-ния погоды, что обусловливает и разницу в сроках начала лёта мушек то отдельным районам Советского Союза.
Под Ленинградом начало лёта отмечено 29 мая — 13 июня. За 17 лет наблюдений в окрестностях Москвы (К. Э. Линдеман, М. К. Шестериков, А. Д. Баранов, Е. А. Покровский, И. М. Беляев) появление шведской мухи на полях установлено в сроки между 3 мая и 8 июня, чаще всего в двадцатых числах мая. Начало лёта мух связано с тремя предшествующими звеньями в ее развитии: 1) возрастом личинок, ушедших на зимовку; 2) временем весеннего коконирования и 3) продолжительностью развития куколки. Все эти моменты находятся в тесной зависимости от температуры и влажности. Холодная весна (апрель и май) задерживает начало коконирования, развитие куколки и вылет мушек. В годы позднего появления шведской мухи (первая декада июня) отмечалось похолодание в мае, средняя месячная температура была на 2—3° ниже средней многолетней.
Раннее появление мушек (начало мая) вызывает теплая погода в апреле — на 1—7° выше средней многолетней.
По наблюдениям А. А. Писнячевского, массовый вылет наступает в то время, когда средняя температура почвы на глубине 1 м достигает 8—9°.
Риггерт связывает выход первых мушек с зацветанием одуванчика, а начало массового лёта с зацветанием ранних сортов яблонь. Блюнк отмечает, что массовый лёт шведской мухи происходит обычно в период цветения ранних сортов яблонь, а если предшествующая осень была достаточно теплой, то и несколько раньше этого срока.
По нашим наблюдениям, сроки появления мушек в Московской области больше совпадают с цветением яблони, чем с цветением одуванчика.
По наблюдениям А. И. Шернина, в Кировской области за 3—5 дней до начала лёта шведской мухи зацветает купальница европейская.
Знание суточной активности насекомых представляет практический и теоретический интерес, особенно при составлении прогнозов.
Для изучения суточного режима шведской мухи нами проводились в июне и июле на протяжении 11 дней специальные учеты лёта мушек и яйцекладки. С утра и до вечера через каждый час подсчитывали яйца на 200 растениях в фазе второго листа. Лёт шведской мухи определялся кошением сачком на меже на одной и той же площадке. В назначенный час вначале учитывали метеорологические факторы, температуру на поверхности почвы и в будке, влажность воздуха, ветер, облачность; затем проводили кошение сачком и подсчет яиц на растении. В результате установлено следующее. Шведская муха начинает откладывать яйца утром с 8 часов, иногда с 9—10; в середине дня в 11, 12, 13 и 14 часов происходит максимальная откладка яиц, причем наибольшей силы достигает в период 12—14 часов; с 15—16 часов энергия кладки ослабевает и в 18—19 часов прекращается; с 19 часов и до 7 часов утра откладка яиц не происходит, хотя бы температура почвы и была выше 11°.
Нами также проводились учеты лёта мух кошением сачком через каждые 2 часа, с восхода солнца и до заката. Единичные особи попадались при кошении в 4 часа утра. С нарастанием температуры количество шведок увеличивалось, достигало максимума в час дня, после чего шло на понижение до 10 часов вечера; если росы на растениях не было, мухи, вспугнутые сачком, попадались в него даже в такое позднее время, как 10 часов вечера.
Наблюдения показали, что наибольшая жизнедеятельность шведской мухи бывает в 12—14 часов. Ведущую роль играет температура, которая в течение суток имеет одновершинную кривую, достигающую максимума в час дня.
Некоторое увеличение облачности и силы ветра к середине дня не нарушало дневного ритма яйцекладки и лёта шведской мухи. В противоположность температуре относительная влажность воздуха уменьшалась к середине дня, что, возможно, также влияло на жизнедеятельность шведской мухи.
В отдельные дни наблюдались отклонения в ходе суточного режима, когда максимум яйцекладки проходил в 10—11 часов йли в 4 часа Дня, а в 1 час Дня наблюдалось заметное понижение, что было вызвано изменением метеорологических факторов.
Интересно отметить, что утром (до 8—9 часов) температура часто бывает значительно выше 11°, однако откладка яиц не .происходит; также и вечером (к 6—7 часам) яйцекладка ослабевает, несмотря на то что температура почвы высокая, иногда не меньше 20°. Возможно, что активность светолюбивой шведской мухи в данном случае связана с ослаблением силы солнечного света; ночью яйцекладка прекращается, что обусловливается не только понижением температуры, но главным образом отсутствием света.
Нами в течение 3 лет изучалась с мая по октябрь продолжительность яйцекладки шведской мухи. С этой целью через каждые 5 дней высевали овес на площадках 1X6 м. С появлением у всходов второго листа подсчитывали ежедневно количество отложенных яиц на 300 растениях.
Установлено, что откладка яиц начинается весной вскоре после вылета мух из ложнококонов и продолжается все лето до последних чисел сентября, то повышаясь, то понижаясь, в зависимости от метеорологических факторов и появления яйцекладущих мушек. За весь летний период вполне ясно выделяются два максимума яйцекладки: первый (соответствует первой летней генерации) происходит в июне — в первой декаде июля и второй максимум — во второй— третьей декаде августа (с 12—15 августа по 23 августа — 8 сентября), то есть соответствует осенне-весенней генерации на озимых посевах.
Вскрытие самок на созревание яичников подтвердило правильность определения двух максимумов кладки яиц. В конце мая—-начале июня преобладают самки с недоразвитыми яичниками; месяц спустя, в конце июня— начале июля, наблюдается наибольшее число мушек со зрелыми яичниками; идет максимальная кладка яиц; с середины июля встречаются в большом количестве самки, отложившие все яйца и израсходовавшие весь запас жирового тела. В середине августа снова появляются мушки со зрелыми яичниками.
Ход яйцекладки в природе чрезвычайно подвержен воздействию внешних факторов, особенно температуры.
Наступление интенсивной откладки яиц весной обыкновенно совпадает с наступлением теплой погоды (19— 21°). Отмеченные два максимума как в июне, так и в августе проходят всегда при высокой температуре— 19— 30°. Понижение яйцекладки в летние месяцы при наличии яйцекладущих самок вызвано временным похолоданием, когда температура днем падает до 11 —16°. Срок осеннего максимума и конец яйцекладки полностью зависят от температуры этого периода.
Поздней осенью, с наступлением холодной и дождливой погоды, мухи вымирают.
Зимуют личинки преимущественно третьего (последнего) возраста. В теплую осень некоторая часть личинок закукливается, весной из этих ложнококонов мухи вылетают раньше.
Вопрос о возрасте зимующих личинок весьма важен, с ним связаны выживаемость личинок в период зимовки и срок появления мушек весной. Установлено, что личинки третьего возраста с хорошо развитым жировым телом дружно коконируются весной и за счет них происходит массовый вылет мушек.
Вредоносность шведской мухи. Шведскую муху относят к так называемым физиологическим вредителям, которые не съедают растение полностью, но своим повреждением нарушают его нормальный рост и развитие. Личинка шведской мухи живет внутри нижней, этиолированной части молодого стебля, вблизи узла кущения. Питается нежными эмбриональными тканями, но узел кущения и пазушные почки обыкновенно не повреждает. При вскрытии поврежденного стебля можно видеть, как вся этиолированная часть выедена личинкой.
Повреждение растущих тканей приостанавливает рост стебля: больной стебель остается в той фазе развития, на которой застало его повреждение. Центральный лист вследствие повреждения его нижней части вянет и желтеет. В результате получается характерный тип повреждения молодого стебля: центральный лист желтый, боковые зеленые. Обыкновенно внешняя картина повреждения (пожелтение листа) выявляется на 8—16-й день от начала питания личинки. Иногда поврежденный лист отсутствует: он или отпадает, или не выходит из листового влагалища.
Поврежденный центральный лист останавливается в росте, и по его длине можно определить, в какой момент началось повреждение стебля.
По фазе молодого стебля и длине поврежденного листа можно восстановить, когда было отложено яйцо, то есть когда произошло заражение. Ориентировочно можно считать, что если поврежден стебель в фазе третьего листа, заражение было во втором листе; если поврежден стебель второго листа, заражение произошло в первом листе.
Меры борьбы.
1. Ранний срок посева яровых хлебов и посев озимых в оптимальные, лучшие агротехнические сроки, установленные для каждого района.
2. Комплекс агротехнических мероприятий, способствующий сильному росту и развитию кукурузы, яровых и озимых хлебов, что увеличивает устойчивость растений к шведской мухе. К этим мероприятиям прежде всего относятся хорошая обработка почвы, удобрения, посев лучшими семенами.
3. Глубокая зяблевая вспашка, которая уничтожает оставшиеся в стерне и падалице яйца, личинки и ложно-коконы шведской мухи. Кроме того, яровые хлеба, посеянные по своевременно и хорошо выполненной зяби, лучше развиваются и становятся более выносливыми к повреждению шведской мухой.
4. Высев устойчивых сортов ячменя, яровой пшеницы, овса и кукурузы.
5. Химические меры борьбы. В борьбе со шведской мухой применяют в основном 12%-ный дуст гексахлорана. Попадая на тело насекомых или даже при небольшом контакте через лапки, гексахлоран вызывает отравление. Минут через 10—15 наступает паралич, мушки не могут передвигаться и летать, падают на землю и в парализованном состоянии находятся до полутора суток, а затем погибают.
По нашему определению, численность шведской мухи на посевах снижается после опыливания гексахлораном на 66—93%, а 5,5%-ным дустом ДДТ —на 36—40%.
В нечерноземной зоне при ранних посевах яровых хлебов главные стебли обычно уходят от повреждения шведской мухой. Применение химических средств защиты сохраняет вторичные продуктивные стебли. Поэтому ячмень, яровую пшеницу, овес опыливают в начале кущения, когда главные стебли имеют 4—5 листьев, а вторичные 1—2 листа, расходуя 12%-ного дуста гексахлорана 10—15 кг на 1 га. По нашим учетам, повреждаемость стеблей снижалась после опыливания гексахлораном на 72%, а дустом ДДТ —на 46%. Прибавка урожая яровой пшеницы от опыливания гексахлораном составила 20%.
Белорусский научно-исследовательский институт земледелия (А. Ф. Марковец) применял авиаопыливание ячменя и овса 12%-ным дустом гексахлорана из расчета 8 кг на 1 га. На торфяных почвах получено при опы-ливании 15 ц ячменя и 13 ц овса с гектара, а без опыливания только 5—6 ц.
Опыливать посевы кукурузы нужно в период от появления всходов (шильце) до четырех листьев. Это очень короткий срок — 6—8 дней.
Имеются рекомендации проводить два опыливания: первое в фазу 1—2 листьев и второе через 5—6 дней. Наш семилетний опыт показал, что достаточно одного опыливания, поскольку второе приходится на время появления четвертого листа и оно уже малоэффективно.
Опыливание кукурузы в фазе первого-третьего листа 12%-JHbrM дустом гексахлорана при норме 10—15 кг на 1 га мы проводили на производственных посевах в Научно-исследовательском институте сельского хозяйства нечерноземной зоны в течение 7 лет (1955—1961). Повреждаемость растений снижалась с 24—40% до 2—11%.
В Калужской области при однократном опыливании гексахлораном повреждаемость кукурузы снизилась с 53,4 до 6,2% (В. М. Тарасевич), в Горьковской области—с 51,9—56,7 до 4,4—5,2% (Я. Н. Харитонов).
Для опрыскивания очень важно подобрать такой препарат, чтобы он обладал высокой токсичностью против злаковых мух, хорошей удерживаемостью на растениях, длительностью действия и небольшой стоимостью. Из современных препаратов, выпускаемых химической промышленностью, нами испытывались против шведской мухи следующие: эмульсия ДДТ, 12%-ный дуст гексахлорана, тиофос, хлорофос, рогор и др. Лучшие результаты получены при опрыскивании посевов 3—4%-ной суспензией гексахлорана, расходуя на гектар 9—12 кг препарата. Повреждаемость стеблей шведской мухой снизилась у яровой пшеницы, овса, ячменя с 22—30% (контроль) до 3—7%, ОТ хлорофоса (0,2%) До 8—10%, тиофоса (0,1%) до 11%, ротора (0,2%) до 8-9%. С уменьшением числа поврежденных стеблей увеличивается продуктивная кустистость на 13—30%.
В 1954 г. нами проводилось испытание 20%-ной ми-неральномасляной эмульсии ДДТ. Опрыскивание яровой пшеницы 0,3%-ной эмульсией снизило повреждаемость стеблей с 34 до 4,3%. Было отмечено, что увеличение концентрации эмульсии до 0,5% вызывало угнетение всходов ржи — листья имели бледно-зеленую окраску.
На протяжении ряда лет нами проводилось на полях Научно-исследовательского института сельского хозяйства нечерноземной зоны комплексное опрыскивание посевов яровых хлебов преимущественно 3—4%-ной суспензией гексахлорана с добавлением соответствующего количества гербицида.
В 1961 г. опрыскивание проводили 5 июня в начале кущения яровой пшеницы, главный стебель имел 4 листа. Сорняки в это время были еще мелкие и слабые. На гектар расходовалось в среднем 250 л воды, 10 кг 12%-ного дуста гексахлорана и 600 г бутилового эфира.
Через 5 дней после опрыскивания яровой пшеницы проведенный учет показал резкое снижение (в 5—6 раз) численности полосатой хлебной блохи и шведской мухи. Этим была достигнута хорошая защита посевов от шведской мухи; отмечено всего лишь 2,5% поврежденных стеблей (в контроле 30%). В течение вегетации пшеница стояла чистая, свободная от сорняков. Количество колосоносных стеблей в посеве увеличилось на 28—34%.
В 1963 г. комплексное опрыскивание ячменя проведено 29—31 мая при помощи опрыскивателя ОСШ-15А аминной солью (1 кг на 1 га) и 12%-ным гексахлораном (10 кг на 1 га). Ячмень был в фазе 3—4 листа, начале кущения; сорняки мелкие. Учетами в период колошения установлено значительное снижение повреждаемости стеблей шведской мухой — с 19 (контроль) до 3%.
Перспективно комплексное опрыскивание посевов кукурузы против сорняков и шведской мухи. В 1960 г. мы впервые провели производственное испытание этого метода. При помощи конного опрыскивателя обработали 7—8 июня 15 га кукурузы в фазе 2—3 листа. Расхо-колошения ячменя. Обработка посевов Ю%-ным масляным раствором ДДТ или 3%-ным раствором гексахлорана проводилась аэрозольным генератором АГ-Л6 при высокой численности шведской мухи, когда на 100 взмахов сачком улавливалось 50—60 мух. На гектар расходовали 10—15 л раствора. Действие препаратов продолжалось до 7 дней, а наиболее эффективно в первые 2— 3 дня после обработки.
В Советском Союзе аэрозоли применил в 1960 г. Ю. П. Александров на яровой пшенице, посеянной на полях орошения. Весной при температуре 16° 8%-ный раствор ДДТ в дизельном топливе (5 л на 1 га) был применен против первого поколения шведской мухи. Перед обработкой улавливалось на 100 взмахов сачком не менее 80 мух, а через 8 часов после применения аэрозолей встречались единичные экземпляры.
В день обрабатывали 300 га пшеницы. Однократная обработка позволила снизить повреждаемость стеблей с 27 (контроль) до 15%, а двукратная — до 5%.
В борьбе со шведской мухой можно также использовать свойство мух питаться нектаром цветов различных культурных и сорных растений, подобрав соответствующую приманку с добавлением инсектицида.
Полевые опыты показали, что опрыскивание яровых хлебов в период кущения 1%-ным раствором рафинадной патоки или глюкозы с добавлением 0,5% кремне-фтористого натрия снижает численность мух на 90%.
Шведская муха обладает хорошо развитым обонянием. Нами были испытаны различные ароматические вещества, которые добавлялись к патоке для усиления ее действия. В результате было установлено, что амилацетат, терпинеол, эфирные масла (анисовое, фенхель-ное, мятное и др.) привлекают мух и добавление этих веществ в небольшом количестве (0,01%) повышает эффективность приманки.
В борьбе с мухами применение сладких отравленных приманок имеет то преимущество, что опрыскивание проводится не сплошное, а в виде приманочных полос, что значительно сокращает расход жидкости на гектар и увеличивает производительность работ.
В совхозе «Раменское» Московской области посевы овса опрыскивали из конного опрыскивателя полосами по ширине захвата опрыскивателя, оставляя неопрысну-тые промежутки в 10 м. На гектар расходовалось 50 Л жидкости. Смертность мух достигала 95%; повреждае.-мость стеблей уменьшалась в 3 раза — с 39 (контроль) до 12%.
Шведскую муху отпугивают такие вещества, как нафталин, парадихлорбензол, гербициды и др. Добавление их к ДДТ, гексахлорану и т. п. увеличивает эффективность ядохимикатов в борьбе со шведской мухой.
Для борьбы с личинками шведской мухи весьма перспективно лечение растений препаратами, проникающими в растительные клетки, или, как их называют, системными инсектицидами. Опыты показали, что чем больше содержится в гексахлоране гамма-изомера, тем сильнее внутрирастительное действие препарата на вредных насекомых. ДДТ также проникает в ткани растений, но его внутрирастительное действие (в частности, на шведскую муху) вдвое слабее гексахлорана.
Мы протравливали семена меркураном, содержащим 12 и 24% гамма-изомера гексахлорана. Проведя ряд опытов, мы установили явное снижение повреждаемости главных стеблей личинками шведской мухи на яровых и озимых посевах.
При посеве семенами, обработанными меркураном; находящийся в нем гамма-изомер, проникая впоследствии в листья (особенно в первые), отравляет насекомых, питающихся этими листьями. С дальнейшим развитием растения и увеличением количества листьев остается в растительных тканях все меньше гамма-изомера. В созревающем зерне его вообще не бывает, что подтвердил проведенный нами специальный опыт с амбарными долгоносиками.
Наши исследования, проведенные на протяжении ряда лет, показали, что увеличение содержания гамма-изомера гексахлорана в протравителе семян до 40% повышает эффективность борьбы с личинками шведской мухи и других скрытостебельных вредителей.
Повреждаемость главных стеблей шведской мухой снизилась на 36% при содержании в меркуране 14% гамма-изомера гексахлорана, а с увеличением его в препарате до 42% повреждаемость снизилась на 58%. При дальнейшем увеличении гамма-изомера до 68% мы не получили более существенного понижения повреждаемости главных стеблей.
Таким образом, протравливание семян комбинированными препаратами, имеющими повышенное содержание гамма-изомера гексахлорана, в некоторой степени (на 50%) предохраняет всходы от повреждения шведской мухой.
Защита зерновых культур - Шведская муха